

배부르게 저녁 식사를 마친 후, 눈앞에 놓인 초콜릿 케이크를 보고 군침을 삼켜본 경험이 있으신가요? 분명 몸은 더 이상 에너지가 필요 없다고 말하는데, 왜 우리는 그 달콤한 유혹을 뿌리치기 어려울까요?
이 질문은 우리 뇌 속에 식욕을 관장하는 두 가지의 서로 다른, 그러나 긴밀하게 연결된 시스템이 존재함을 암시합니다. 하나는 생존을 위해 에너지 균형을 맞추는 '항상성 식욕' 시스템이고, 다른 하나는 에너지 필요와 무관하게 쾌락과 보상을 좇는 '비항상성 식욕' 시스템입니다.
수십 년간 과학자들은 이 두 시스템을 별개로 여겨왔지만, 최신 신경과학 연구는 이들이 어떻게 서로 영향을 주고받으며 우리의 식사 행동을 조종하는지 그 정교한 메커니즘을 밝혀내고 있습니다.
이 글에선 Experimental & Molecular Medicine.논문에 실린 Ahn BH외의 "Brain circuits for promoting homeostatic and non-homeostatic appetite" 리뷰 논문을 바탕으로, 우리 뇌 속 깊은 곳으로 들어가 식욕의 비밀을 파헤쳐 보겠습니다.

생존을 위한 온도조절장치: 항상성 식욕 회로
우리 몸은 정교한 에너지 '온도조절장치'를 가지고 있습니다. 에너지가 부족하면 경고등이 켜져 음식을 찾게 하고, 에너지가 충분하면 식욕을 억제합니다. 이 시스템의 핵심 조절자는 지방세포에서 분비되는 '포만 호르몬'인 `leptin`과 위에서 분비되는 '공복 호르몬'인 `ghrelin`입니다.
이 호르몬들은 혈액을 타고 뇌의 시상하부(hypothalamus)로 이동해 에너지 상태를 보고합니다. 시상하부의 궁상핵(arcuate nucleus, ARC)에는 식욕의 가속 페달과 브레이크 역할을 하는 두 종류의 신경세포가 존재합니다. 바로 `AgRP` 뉴런과 `POMC` 뉴런입니다.

가속과 감속의 길항작용: AgRP와 POMC 뉴런
`AgRP` 뉴런은 식욕의 '가속 페달'입니다. 공복 상태가 되어 `ghrelin` 수치가 높아지면 `AgRP` 뉴런이 강력하게 활성화되어 '먹어라!'라는 신호를 뇌 전체에 보냅니다. 반면, `POMC` 뉴런은 식욕의 '브레이크' 역할을 합니다. 식사 후 `leptin` 수치가 상승하면 `POMC` 뉴런이 활성화되어 포만감을 느끼게 하죠. 이 두 뉴런 그룹은 마치 디지털 회로의 '플립-플롭(flip-flop)' 스위치처럼 작동합니다.
`ghrelin`이 스위치를 'ON'으로 돌리면 `AgRP` 뉴런이 활성화되면서 동시에 `GABA`라는 신경전달물질을 분비해 `POMC` 뉴런의 활동을 억제합니다. 반대로 `leptin`은 이 스위치를 'OFF'로 되돌리려 하죠. 이처럼 정교한 상호 억제 메커니즘을 통해 우리 뇌는 배고픔 또는 배부름 상태를 안정적으로 유지합니다.
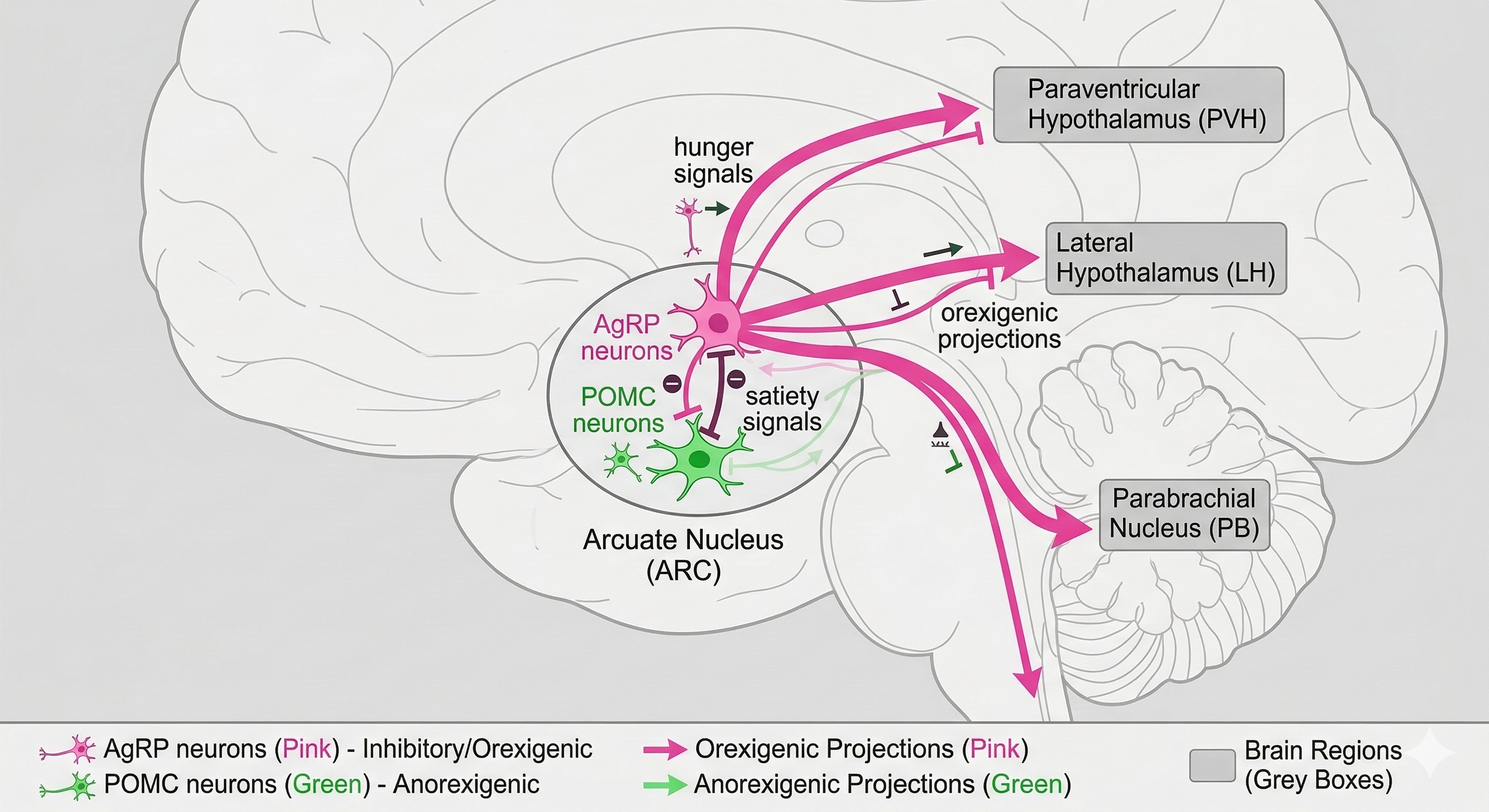
그림 1
그림 1의 회로도는 이 시스템의 복잡한 연결망을 보여줍니다. 중앙 허브인 궁상핵(ARC)에서 분홍색 `AgRP` 뉴런은 초록색 `POMC` 뉴런을 억제하는 동시에, 시상하부의 다른 영역(PVH, LH)과 뇌간(PB) 등 뇌의 여러 지역으로 '배고픔' 신호를 광범위하게 전송합니다.
이 외에도 뇌간의 고립로핵(NTS)은 미주신경절(NG)을 통해 위장의 영양 상태 정보를 받아들이고, 이 정보를 다시 시상하부로 전달하는 등 여러 회로가 유기적으로 협력하여 우리 몸의 에너지 균형을 지킵니다.

쾌락 추구자: 비항상성 식욕 회로
항상성 시스템이 생존을 위한 것이라면, 비항상성 시스템은 '즐거움'을 위한 것입니다. 맛있는 음식의 모습, 냄새, 혹은 그 음식을 먹었던 즐거운 기억만으로도 이 회로는 작동합니다.
이 시스템의 핵심 주자는 시상하부 외측(lateral hypothalamus, LH)에 위치한 `Vgat` 뉴런들입니다. 이 뉴런들은 에너지 결핍 상태가 아닐 때도 활성화되어 음식을 포함한 다양한 보상적 대상에 대한 강력한 섭취 행동을 유발합니다.
심지어 영양가 없는 물건을 씹는 행동까지 유도할 정도입니다. 이들은 단순히 '먹어라'는 신호를 넘어 '저것을 원한다!'는 강력한 동기를 부여하는 역할을 합니다.
보상 회로를 장악하다: 도파민과의 연결
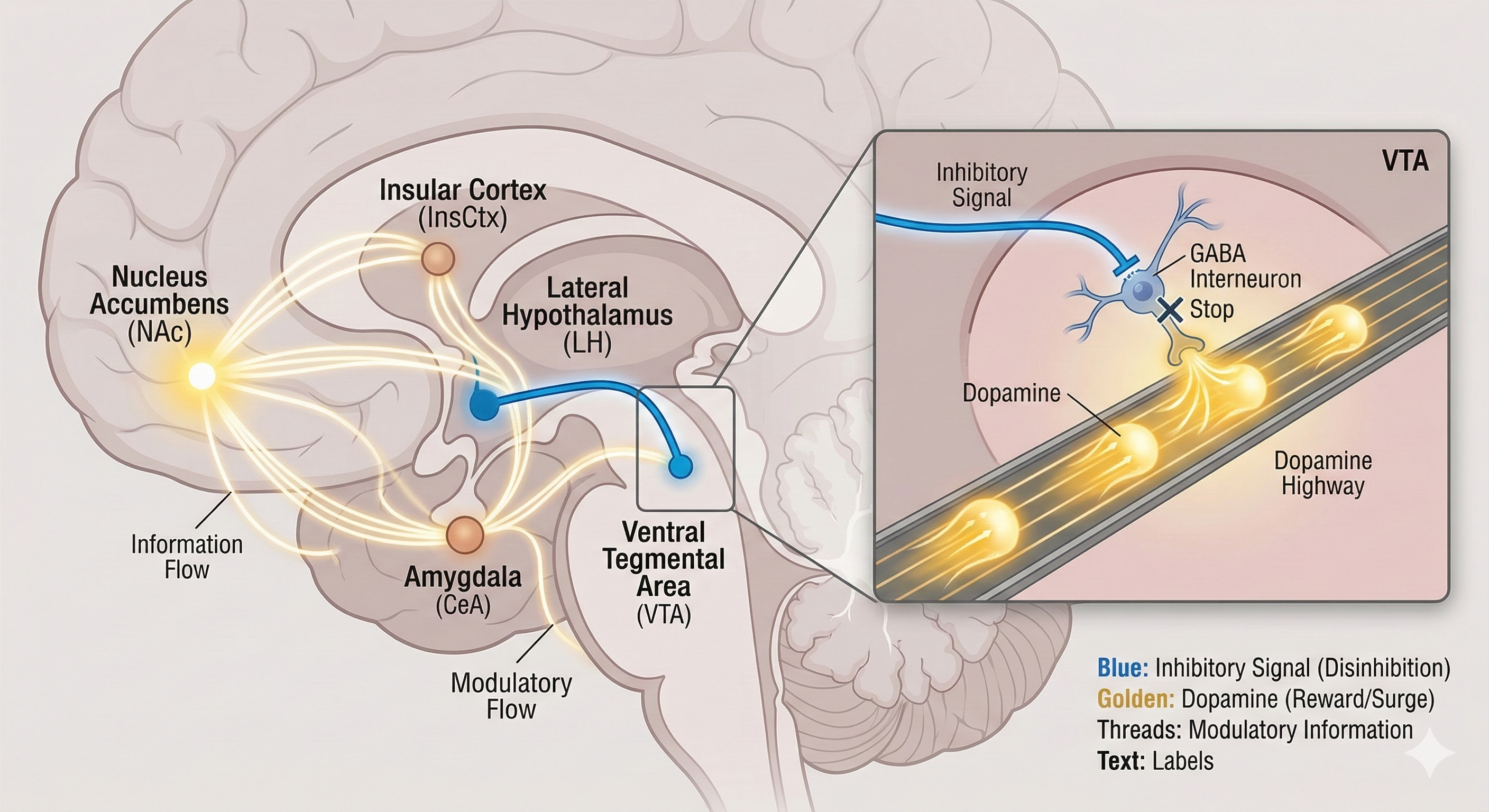
그림 2
그림 2는 이 쾌락 추구 회로의 복잡한 연결망을 보여주는 지도와 같습니다. 핵심 경로는 시상하부 외측(LH)에서 시작됩니다. 여기서 출발한 억제 신호(파란색 선)는 복측피개영역(VTA)으로 향하여, VTA 내의 또 다른 억제 뉴런의 활동을 멈추게 합니다.
이는 마치 브레이크를 밟고 있던 발을 치우게 만들어, 도파민이라는 자동차가 측좌핵(NAc)을 향해 질주하도록 하는 '탈억제' 과정입니다. 이 외에도 뇌섬엽 피질(InsCtx)이나 편도체(CeA) 등 여러 뇌 영역들이 이 네트워크에 복잡하게 얽혀, 음식에 대한 기억과 감정을 강력한 갈망으로 전환시키는 데 기여합니다.
두 세계의 충돌과 융합: 식욕 회로의 상호작용
최신 연구의 가장 흥미로운 발견은 이 두 식욕 시스템이 결코 독립적이지 않다는 것입니다. 배가 고플 때(항상성 신호) 맛있는 음식이 더욱 간절하게 느껴지는(비항상성 반응) 것은 바로 이 두 시스템이 서로 긴밀하게 상호작용하기 때문입니다.
예를 들어, 공복 호르몬 `ghrelin`이나 `AgRP` 뉴런의 활성화는 VTA의 도파민 반응을 증폭시켜 음식의 보상 가치를 높입니다. 즉, 생존 시스템이 쾌락 시스템의 볼륨을 키우는 셈입니다. 반대로, 도파민 신호가 `leptin`의 식욕 억제 효과에 영향을 미치는 등 쾌락 시스템이 생존 시스템을 조절하기도 합니다.
하나의 뉴런, 두 개의 임무
두 시스템의 융합을 가장 명확하게 보여주는 예는 결절핵(tuberal nucleus)의 `Sst` 뉴런입니다. 이 뉴런들은 `ghrelin`에 반응하여 공복 시 일반 사료 섭취를 촉진하는 항상성 역할을 수행합니다. 동시에, 이 뉴런들은 과거에 고지방 식사와 같은 맛있는 음식을 먹었던 장소에 다시 갔을 때 발생하는 '상황 의존적 과식'에도 필수적인 것으로 밝혀졌습니다.
이는 하나의 신경세포 집단이 생존을 위한 배고픔과 쾌락을 위한 갈망이라는 두 가지 임무를 모두 수행할 수 있음을 보여주는 놀라운 사례이며, 두 식욕 시스템이 얼마나 깊이 얽혀 있는지를 증명합니다.
결론
우리의 식욕은 단순히 배를 채우기 위한 단일한 욕구가 아닙니다. 그것은 생존을 위해 에너지 균형을 맞추려는 정교한 '항상성 회로'와, 쾌락과 보상을 추구하며 학습된 기억에 따라 움직이는 강력한 '비항상성 회로' 사이의 끊임없는 대화와 협상의 산물입니다.
현대 사회는 고칼로리의 맛있는 음식이 넘쳐나기 때문에, 우리의 쾌락 추구 시스템이 생존 시스템을 압도하기 쉬운 환경입니다. 비만이나 섭식 장애와 같은 질병의 근원을 이해하기 위해서는 이 두 시스템의 상호작용을 파헤치는 것이 중요합니다. 신경과학의 발전은 이 복잡한 회로 지도를 더욱 정밀하게 그려내고 있으며, 이는 미래에 식욕 관련 질환에 대한 새로운 치료법을 여는 열쇠가 될 것입니다.
정리한 논문 : Ahn BH, Kim M, Kim S-Y. Brain circuits for promoting homeostatic and non-homeostatic appetites. Experimental & Molecular Medicine. 2022;54:349-357. doi:10.1038/s12276-022-00758-4