

20세기 생물학을 지배했던 오래된 패러다임 중 하나는 단백질이 활성(ON) 또는 비활성(OFF)이라는 단 두 가지의 고정된 상태로만 존재한다는 것이었습니다. 이를 바탕으로 신약 개발은 약물이 단백질에 결합할 때 형태가 빠르게 맞춰진다는 '유도 적합(induced fit)' 모델에 크게 의존해 왔습니다.
하지만 생물물리학의 발전과 분자동역학(MD) 시뮬레이션은 단백질의 자유 에너지 지형(free energy landscape)이 훨씬 복잡하다는 것을 보여줍니다. 단백질은 나노초(ns)에서 밀리초(ms)에 이르는 동역학적 장벽(kinetic barrier)을 넘나들며 끊임없이 움직이고, 다양한 구조적 상태가 동적 평형을 이루는 '구조적 앙상블(conformational ensembles)'을 형성합니다.
이러한 역동적인 관점의 전환은 약물이 미리 존재하는 특정 구조를 선택해 결합한다는 '구조 선택(conformational selection)' 모델을 대두시켰으며, 기존에는 공략할 수 없었던 단백질의 숨겨진 주머니(cryptic pocket)를 표적으로 삼는 '알로스테릭(allosteric)' 신약 개발의 새로운 지평을 열고 있습니다.
결합력(Affinity)을 넘어 효능(Efficacy)으로
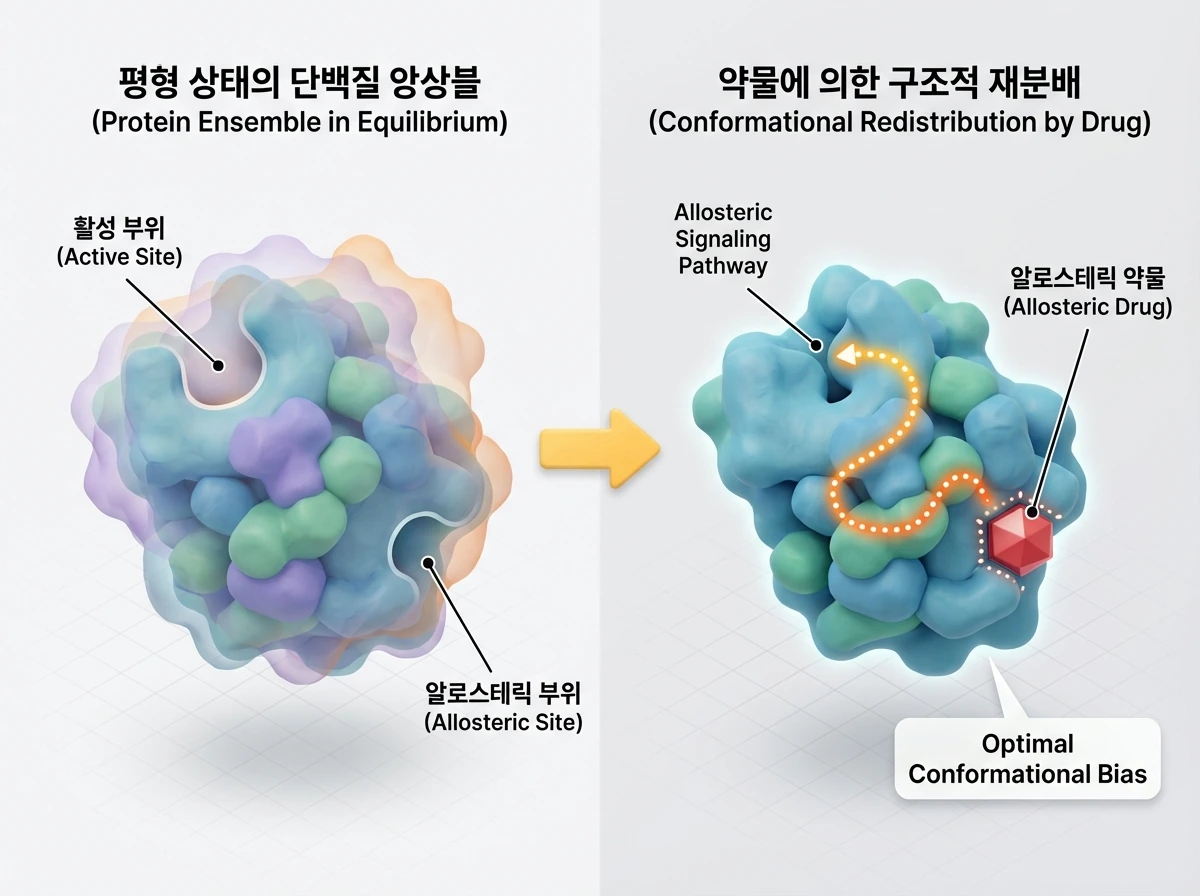
전통적인 신약 설계에서는 약물이 표적에 얼마나 단단히 결합하는지를 나타내는 친화도(affinity)가 가장 중요했습니다. 하지만 알로스테릭 약물 개발에서는 친화도 못지않게 '효능(efficacy)'이 핵심적인 역할을 합니다. 여기서 효능이란 약물이 단백질의 특정 구조적 상태를 안정화하여 전체 앙상블의 평형을 원하는 방향으로 이동시키는 '구조적 재분배(conformational redistribution)' 능력을 말합니다.
흥미롭게도 약물과 단백질 포켓 사이의 미세한 기하학적, 정전기적 불일치가 오히려 국소적인 구조 변화를 유도하고, 이 신호가 활성 부위로 전달되어 강력한 알로스테릭 효과를 냅니다. 즉, 완벽하게 들어맞는 것보다 표적의 '최적의 구조적 편향(optimal conformational bias)'을 영리하게 유도하고 알로스테릭 신호 전달 경로를 매핑(mapping)하는 것이 치료 목적 달성에 더 중요해진 것입니다.
mTOR 억제제의 진화: 두 마리 토끼를 잡는 이중 표적 전략
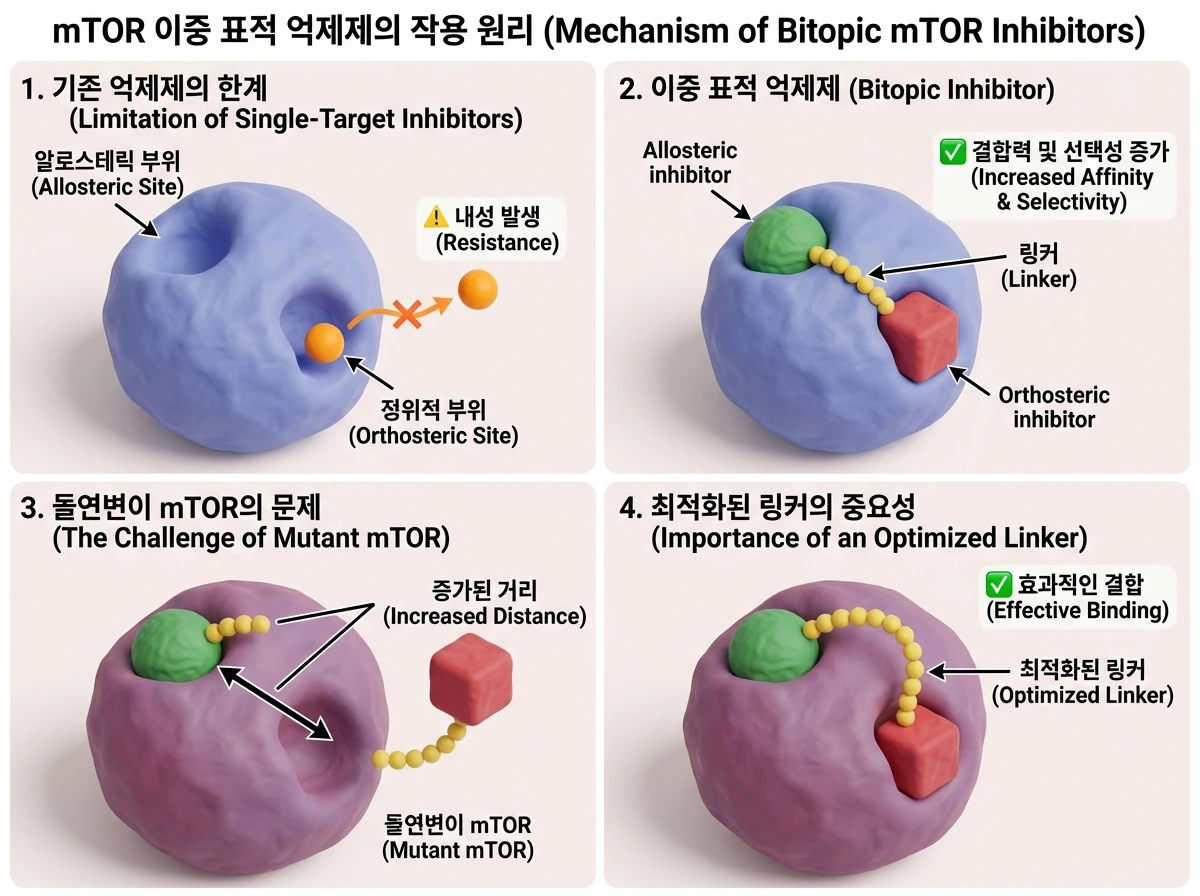
세포 성장과 대사를 조절하는 핵심 인자인 mTOR는 항암제 개발의 주요 표적입니다. 하지만 기존의 알로스테릭 약물(예: everolimus)이나 정위적(orthosteric) 억제제를 단독으로 사용할 경우, 단백질의 구조적 이질성(heterogeneity)이 증가하며 내성 돌연변이가 빈번하게 발생했습니다.
이를 극복하기 위해 등장한 것이 두 부위를 동시에 공략하는 이중 표적(bitopic) 억제제입니다. 예를 들어, Rapalink-1이나 RMC-5552 같은 약물은 알로스테릭 억제제와 정위적 억제제를 링커(linker)로 연결하여 결합력과 선택성을 극대화하고 약물의 체류 시간(residence time)을 연장합니다. 최근 연구에 따르면, 발암성 돌연변이를 지닌 mTOR는 활성 부위가 열린 상태로 존재하는 비율이 높으며, 이로 인해 두 결합 부위 간의 거리가 멀어질 수 있습니다.
따라서 돌연변이의 고유한 구조적 앙상블을 이해하고 두 결합 부위 사이의 거리를 좁힐 수 있도록 링커의 길이를 최적화하는 것이 약물의 효능을 결정짓는 중요한 변수가 되며, 지나치게 긴 링커가 유발할 수 있는 독성 문제라는 한계점 역시 함께 고려되어야 합니다.
EGFR 돌연변이의 두 얼굴: 폐암과 뇌종양의 차이
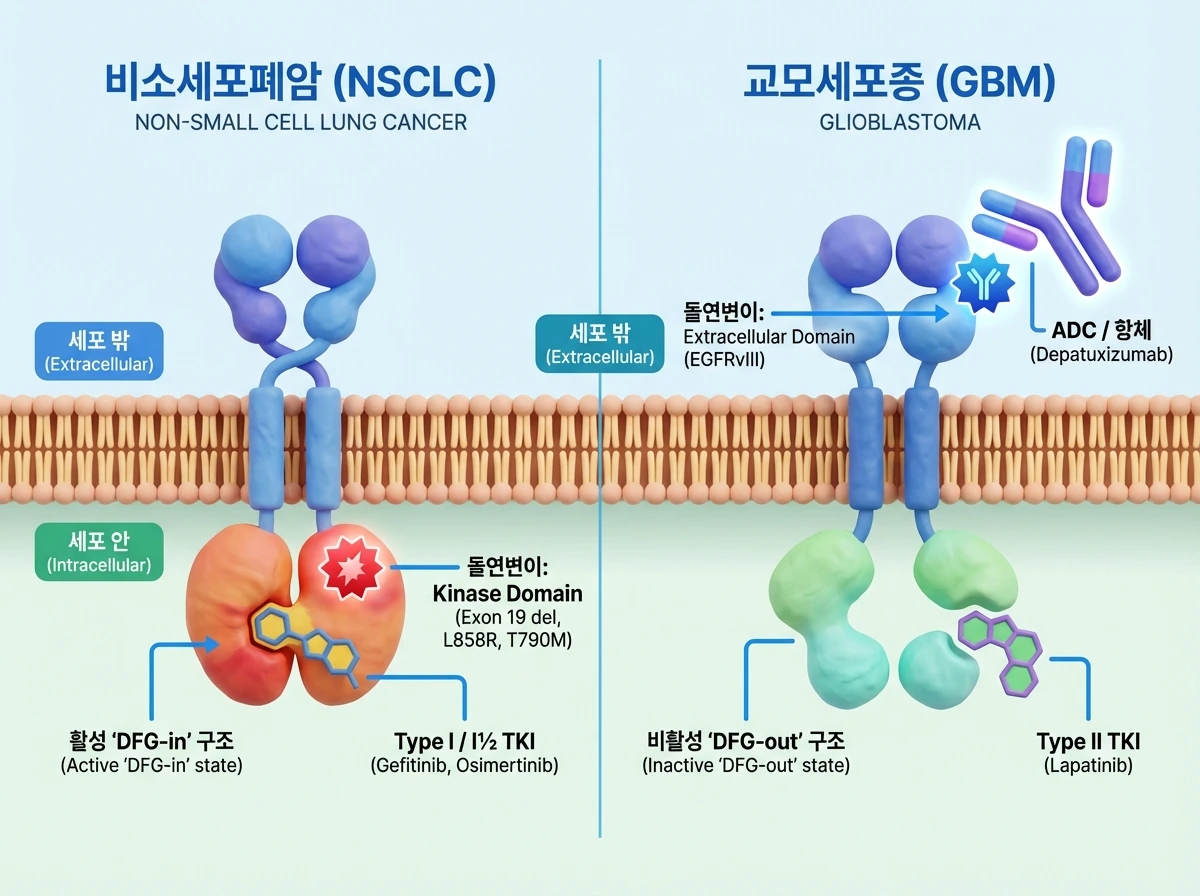
EGFR은 암종에 따라 돌연변이가 발생하는 위치와 약물에 대한 반응성이 완전히 다릅니다. 비소세포폐암(NSCLC)에서는 주로 세포 내 인산화효소 도메인(kinase domain)에 돌연변이(예: 엑손 19 결손, L858R, T790M)가 생깁니다. 이 경우 수용체는 비대칭적(asymmetric) 이합체를 형성하며 활성화된 'DFG-in' 구조를 띠게 되는데, 여기에 결합하는 제1형(Type I) 정위적 티로신 키나아제 억제제(TKI)인 Gefitinib이나 Erlotinib이 효과적입니다. 또한, T790M 내성 돌연변이를 극복하기 위해 개발된 3세대 비가역적 억제제 Osimertinib은 C797 잔기와 상호작용하며 활성 상태의 인산화효소 도메인에 결합하는 제1½형(Type I½) 억제제로 작용합니다.
반면, 교모세포종(GBM)에서는 세포 밖 외부 도메인(extracellular domain)에 주로 돌연변이(예: R108K, A289X, G598X 및 엑손 2-7이 결손된 EGFRvIII)가 발생합니다. 흥미롭게도 이러한 세포 밖의 돌연변이는 단백질 앙상블의 변화를 유도하여 세포 내 도메인을 대칭적(symmetric) 이합체와 유사한 비활성 상태로 알로스테릭하게 조절합니다. 그 결과, GBM에서는 비활성 'DFG-out' 구조에 결합하는 제2형(Type II) TKI인 Lapatinib이나, 돌연변이가 생긴 외부 도메인을 직접 표적으로 삼는 항체-약물 접합체(ADC)인 Depatuxizumab이 효과를 발휘합니다.
더불어, 세포 밖 외부 도메인을 겨냥하는 전략으로는 Matuzumab이나 Panitumumab 같은 단일클론항체도 있습니다. 이들은 리간드 결합이나 수용체의 이합체화를 물리적으로 차단하여 EGFR의 활성화를 막습니다. 이처럼 종양의 미세환경과 돌연변이 위치에 따라 단백질이 선호하는 구조적 상태(비대칭적 활성 상태 vs 대칭적 비활성 상태)가 어떻게 달라지는지 파악하는 것은 맞춤형 치료제 개발에 필수적입니다.
융합 단백질의 저항성과 엑소사이트(Exosite)의 잠재력
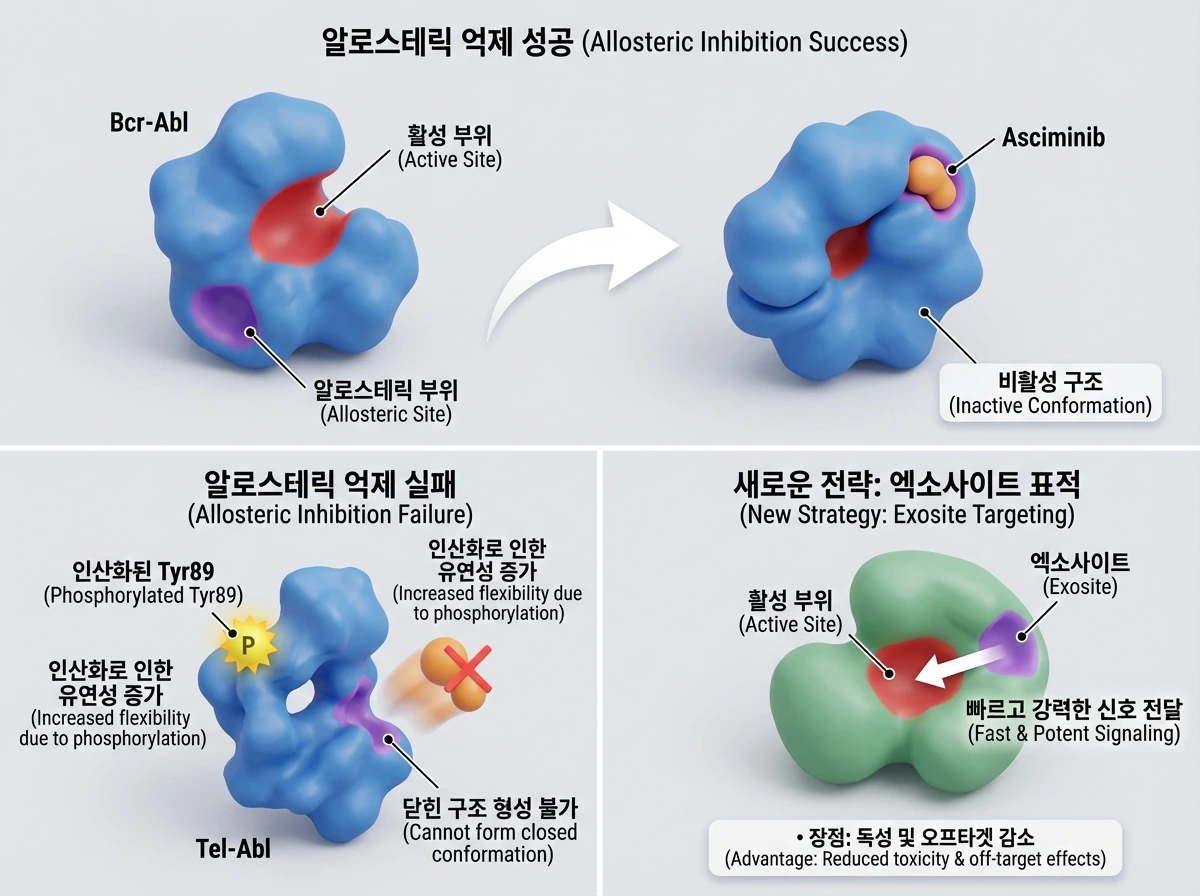
백혈병을 유발하는 Abl 융합 단백질의 사례는 표적 특이적인 활성화 기전(target-specific activation mechanisms)을 이해하는 것이 얼마나 중요한지 보여줍니다. Bcr-Abl은 알로스테릭 억제제인 asciminib에 잘 반응하지만, Tel-Abl은 그렇지 않습니다. Tel-Abl의 특정 잔기(Tyr89)에 인산화가 일어나면 단백질의 유연성이 증가하여 억제제가 결합할 수 있는 닫힌 구조를 형성하지 못하기 때문입니다.
한편, 활성 부위와 인접한 외부 결합 부위인 '엑소사이트(exosite)'를 표적으로 삼는 전략도 주목받고 있습니다. CSN 단백질 복합체 연구에서 알 수 있듯, 엑소사이트는 활성 부위와의 거리가 짧아 알로스테릭 신호 전달이 빠르고 강력하며, 기존 ATP 경쟁적 억제제가 가지는 독성 및 오프타겟(off-target) 부작용을 줄일 수 있는 매력적인 대안입니다.
결론
결론적으로, 단백질을 고정된 구조가 아닌 역동적인 '구조적 앙상블'로 바라보는 시각은 현대 약리학의 패러다임을 근본적으로 바꾸고 있습니다. 이러한 '앙상블-활성 상관관계(Ensembles-Activity Relationship)'에 기반한 접근은 숨겨진 결합 부위를 찾아내고, 약물의 효능을 정교하게 조절하며, 구조적 이질성으로 인한 내성 돌연변이를 극복하는 데 강력한 무기가 됩니다.
나아가 최근 주목받는 생체분자 응축물(biomolecular condensates)과 같이 밀집되고 제한된 미세환경 내에서 단백질의 앙상블이 어떻게 변화하는지 규명한다면, 표적 단백질 분해(targeted degradation)나 분자 접착제(molecular glues)와 같은 혁신적인 치료 전략은 더욱 정교해질 것입니다. 생명 현상의 역동성을 이해하는 자만이 미래의 신약 개발을 주도할 수 있을 것입니다.