
도입
이 리뷰는 신경세포에서 미토콘드리아를 단순히 ATP를 만드는 소기관이 아니라, 뉴런의 긴 형태와 높은 활동성을 떠받치는 ‘공간적으로 조직된 기능 단위’라는 관점에서 최근 연구 흐름을 정리한다. 뉴런은 축삭이 매우 길고, 활동전위 발생과 이온기울기 회복, 시냅스 가소성 유지까지 동시에 수행해야 하므로 에너지 수요가 크고 그 수요가 세포 전체에 고르게 분산되어 있지 않다. 따라서 어디에 미토콘드리아가 위치하는지, 얼마나 이동하고 멈추는지, 언제 분열·융합하고 제거되는지가 곧 뉴런 기능과 직결된다. 중요한 점은 이 기능들이 서로 따로 존재하지 않는다는 것이다. 같은 미토콘드리아가 축삭에서는 수송의 대상이 되고, 시냅스 근처에서는 ATP와 Ca2+ 조절을 맡으며, 가지점에서는 국소 번역을 돕고, 손상 시에는 품질관리의 표적이 된다. 저자들은 이런 연결성을 바탕으로 미토콘드리아의 위치 조절, ATP 생산, Ca2+ 완충, 신경전달물질 대사, 뉴런-아교세포 대사 협력, 그리고 축삭·수상돌기에서의 국소 단백질 합성까지를 하나의 흐름으로 묶어 설명한다. 따라서 이 리뷰의 핵심은 ‘미토콘드리아가 무엇을 하느냐’보다 ‘어느 구획에서, 어떤 분자 환경 속에서, 다른 세포소기관 및 주변 세포와 어떻게 연결되어 그 기능을 수행하느냐’를 묻는 데 있다.
1. 뉴런에서 미토콘드리아를 따로 봐야 하는 이유
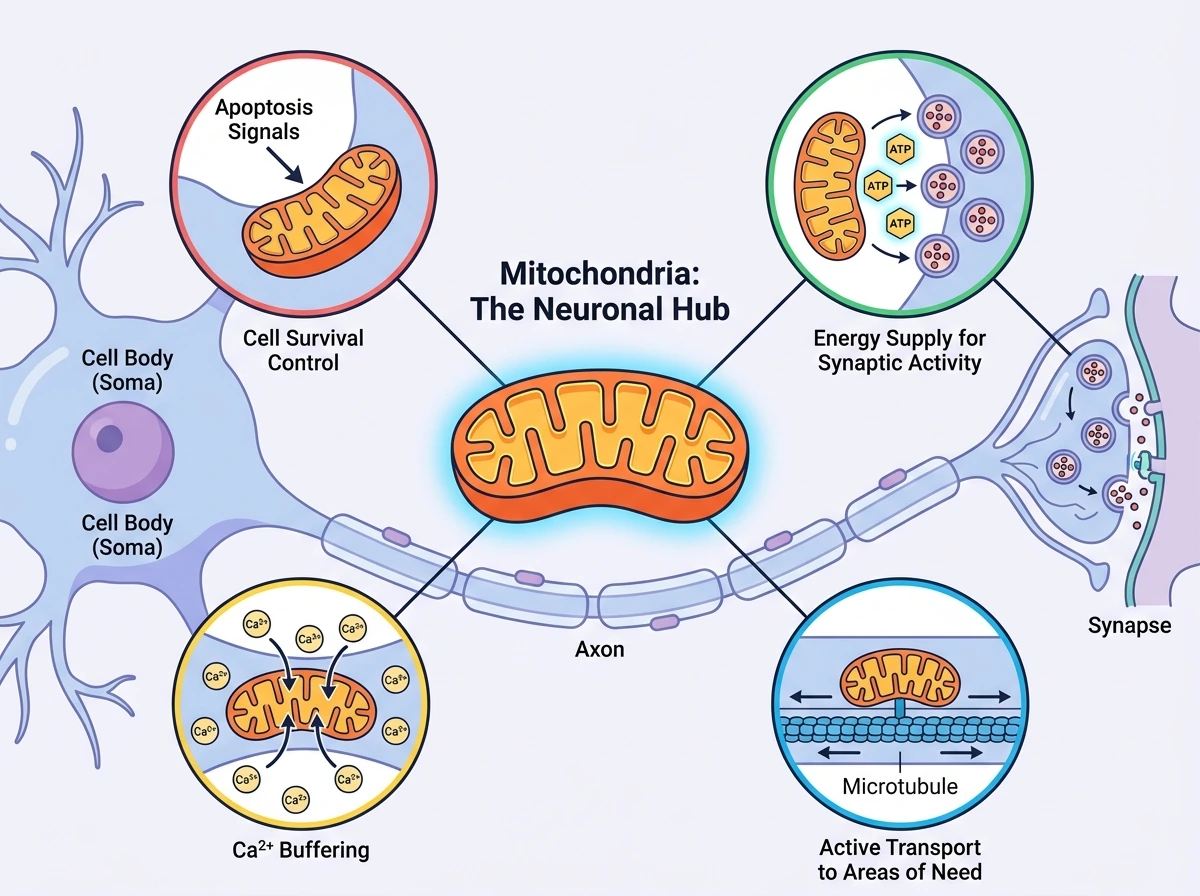 단백질이 합성
단백질이 합성
이 리뷰의 출발점은 뉴런이 일반적인 세포와 다르다는 사실이다. 뉴런은 긴 축삭과 복잡한 수상돌기를 가지며, 정보 전달이 일어나는 위치와 단백질이 합성되는 세포체가 멀리 떨어져 있다. 이런 구조에서는 에너지를 한곳에서만 만들어서는 충분하지 않다. 실제로 세포질의 해당과정은 포도당 1분자당 순수 ATP 2개만을 만들지만, 미토콘드리아의 산화적 인산화는 약 30~32개의 ATP를 생산한다. 따라서 뉴런에서는 미토콘드리아가 단순히 많이 존재하는 것보다, 필요한 자리로 이동해 적절한 시점에 ATP를 공급하는 것이 핵심이다. 더 나아가 미토콘드리아는 Ca2+를 흡수해 세포질 칼슘 급등을 완충하고, TCA 회로를 통해 여러 대사경로와 연결되며, 세포사멸 경로의 관문 역할까지 수행한다. 여기에 더해 뉴런 안에서는 수송, 형태 변화, 국소 번역, 품질관리까지 같은 소기관의 서로 다른 상태로 이어진다. 즉 뉴런에서 미토콘드리아는 에너지 생산, 신호 조절, 생존 결정, 그리고 원거리 구획 유지가 만나는 허브라고 이해하는 것이 이 리뷰의 기본 틀이다.
2. 위치가 기능을 만든다: 뉴런 안에서 미토콘드리아는 어떻게 배치되는가
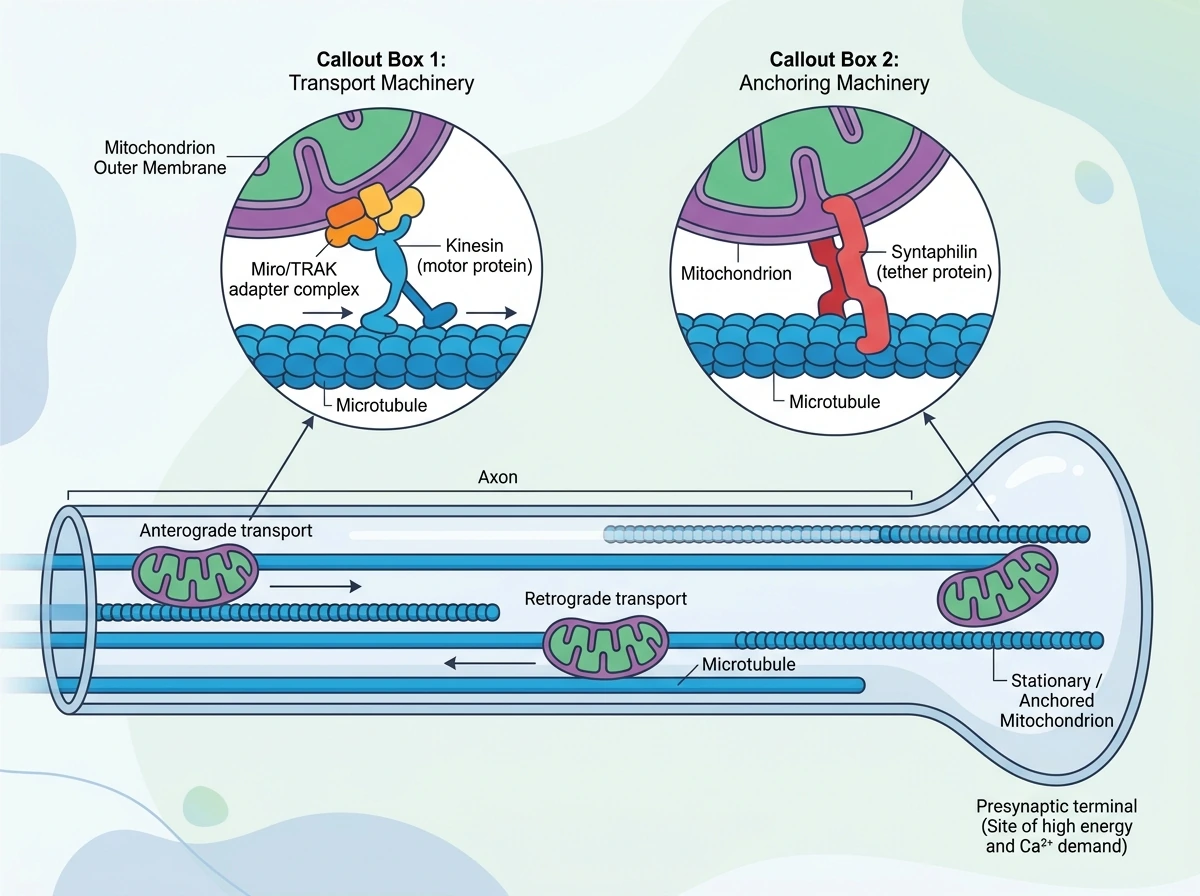
현재 비교적 확립된 견해는 뉴런 기능의 상당 부분이 미토콘드리아의 ‘정확한 위치’에 의존한다는 것이다. 축삭과 수상돌기에서는 미세소관을 따라 장거리 이동이 일어나고, 이때 키네신과 다이네인, 그리고 바깥막 단백질인 Miro와 TRAK 계열 어댑터가 미토콘드리아를 모터 단백질과 연결한다. 따라서 같은 뉴런 안에서도 앞으로 가는 집단과 뒤로 오는 집단이 공존한다. 한편 액틴 세포골격과 미오신은 짧은 거리 이동과 정지를 돕고, syntaphilin은 특히 축삭에서 미토콘드리아를 미세소관에 고정하는 역할을 한다. 흥미로운 점은 모든 미토콘드리아가 계속 움직이지 않는다는 것이다. 실제로는 일부만 이동하고, 많은 미토콘드리아는 잠시 멈추거나 장기적으로 정박한다. 이 정박은 축삭 가지점, 랑비에 결절, 시냅스처럼 에너지나 Ca2+ 조절 수요가 높은 부위에서 두드러진다. 중요한 것은 이런 위치 선택이 단지 ATP 공급 때문만은 아니라는 점이다. 같은 원거리 구획에서 Ca2+ 완충, 소포 순환 지원, 국소 번역 보조가 함께 요구되므로, 정박은 여러 기능을 한 지점에 묶는 방식으로 이해하는 편이 더 정확하다. 정지 메커니즘도 하나가 아니다. Ca2+ 신호, 포도당 유입에 따른 O-GlcNAcylation, 손상에 따른 인산화, syntaphilin 매개 고정, 그리고 신경활동으로 ATP가 떨어질 때 AMP-activated kinase가 작동하여 활성 시냅스 근처로 미토콘드리아를 끌어들이는 경로 등이 함께 작동한다. 즉 ‘어디에 미토콘드리아가 있느냐’는 수동적 결과가 아니라, 활동과 대사 상태에 따라 조절되는 적극적 배치 과정이라는 것이 이 분야의 핵심 합의다.
3. 모든 시냅스에 미토콘드리아가 있지는 않다: 정설과 미해결 문제
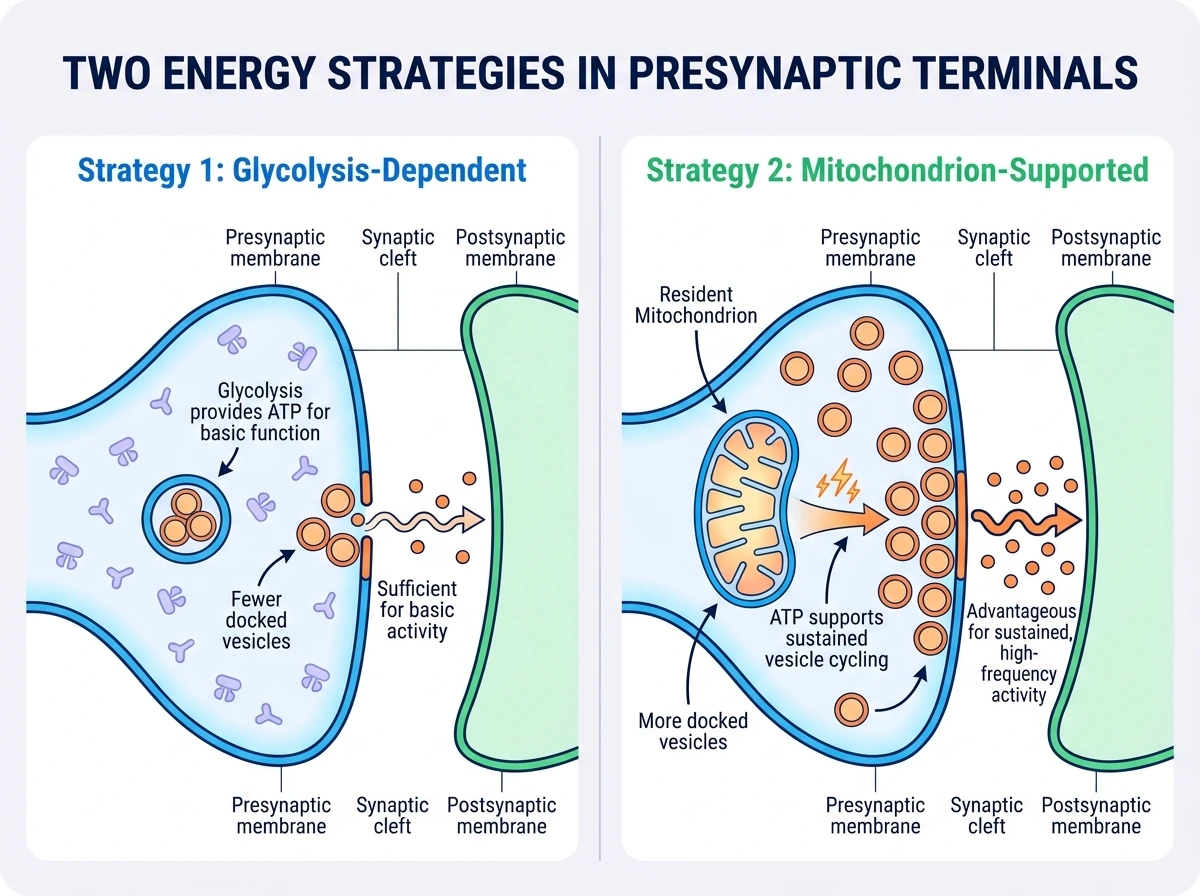
하지만 이 리뷰가 흥미로운 이유는, 이런 위치 조절 원리가 분명해 보이는데도 실제로는 30~50%의 presynapse에 상주 미토콘드리아가 없다는 관찰을 함께 제시하기 때문이다. 이것은 앞 절의 논리와 모순이라기보다, 시냅스가 에너지와 Ca2+ 문제를 푸는 방식이 하나만은 아니라는 뜻에 가깝다. 현재까지의 데이터는 두 가지 점을 함께 시사한다. 첫째, 미토콘드리아가 없는 일부 시냅스에서도 해당과정만으로 기본 ATP 수요를 감당할 수 있다. 실제로 미토콘드리아 유무에 따라 ATP 수준 차이가 뚜렷하지 않았다는 관찰도 소개된다. 저산소 조건에서는 해당과정 효소가 presynaptic bouton에 모여 대사 흐름을 강화할 수 있다는 결과도 이런 적응성을 뒷받침한다. 둘째, 그럼에도 불구하고 지속적이거나 반복적인 시냅스 활동에는 상주 미토콘드리아가 유리하다. 미토콘드리아가 있는 presynapse에서 정박된 시냅스 소포 수가 더 많았다는 결과는, 미토콘드리아 유래 ATP가 장기적·고빈도 활동을 뒷받침한다는 해석과 잘 맞는다. 따라서 현재의 정리는 ‘기본 시냅스 기능에는 항상 미토콘드리아가 필수는 아니지만, 지속적 활동과 안정적 전달에는 현장 배치된 미토콘드리아가 이점이 있다’ 정도다. 왜 일부 시냅스만 미토콘드리아를 갖는지는 아직 미해결이며, 리뷰는 이것이 능동적 배제인지, 축삭으로 들어갈 수 있는 미토콘드리아 수의 한계 때문인지를 앞으로 풀어야 할 문제로 남긴다.
4. 모양과 분포의 논리: 분열과 융합은 왜 중요한가
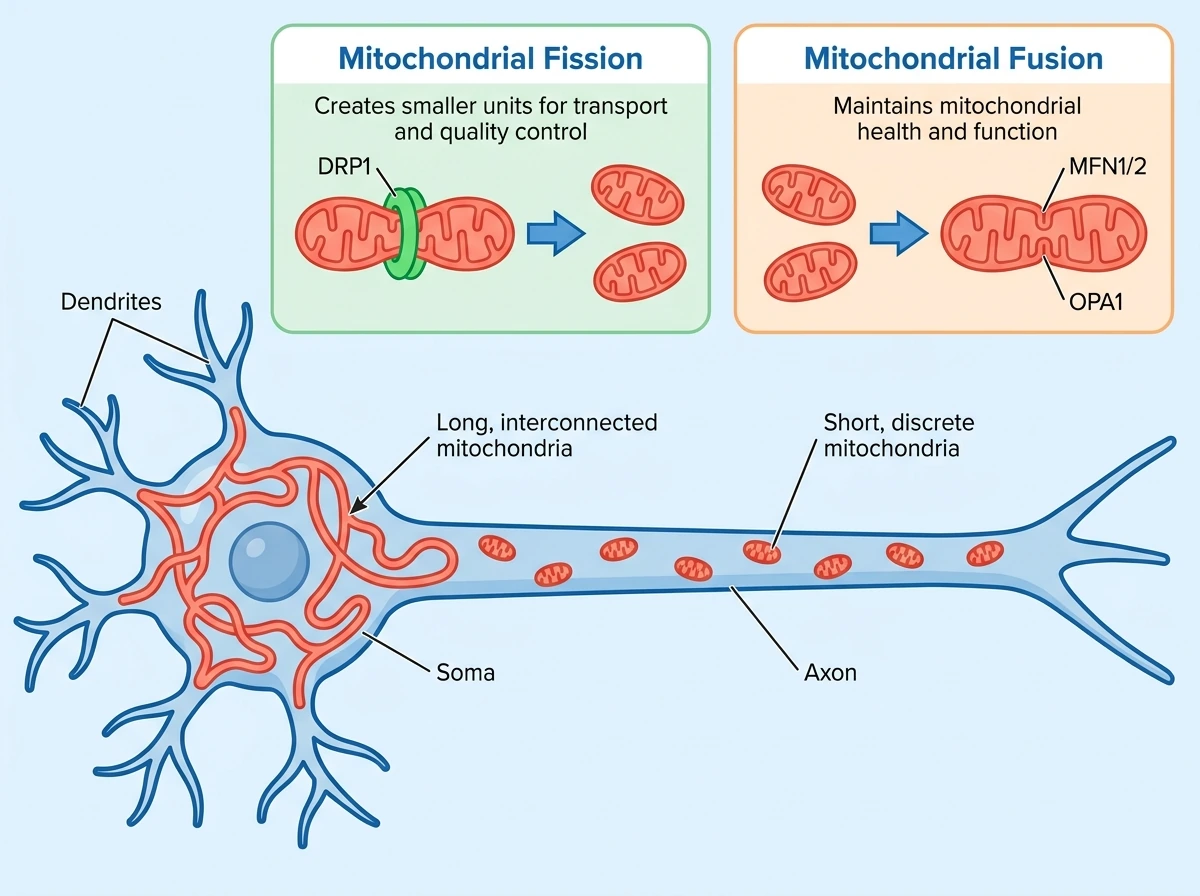
뉴런 내 미토콘드리아는 위치뿐 아니라 형태도 구획별로 다르다. 일반적으로 세포체와 수상돌기 쪽 미토콘드리아는 길고 부분적으로 연결된 관 형태가 많고, 축삭의 미토콘드리아는 더 짧고 비교적 균일하다. 리뷰는 이 형태 차이가 기능 차이를 반영할 가능성을 제기한다. 여기서 분열과 융합은 단지 ‘모양 바꾸기’가 아니라, 배치와 품질관리까지 연결하는 과정으로 이해해야 한다. 미토콘드리아 분열은 작은 단위를 만들어 재배치와 품질관리에 유리하게 한다. 이 과정은 DRP1이 중심이 되며, ER와 액틴이 협력한다. 특히 midzone fission은 생합성과 연관되고, peripheral fission은 작은 조각을 떼어내 품질관리와 mitophagy로 이어지기 쉽다고 정리된다. 반대로 융합은 미토콘드리아 건강과 막전위 유지에 중요하며, 바깥막의 MFN1/2와 안쪽막의 OPA1이 이를 매개한다. 반복 자극 중 시냅스 전달 유지에 융합이 중요하다는 점은 비교적 강한 합의로 제시된다. 다만 ‘축삭 미토콘드리아가 짧은 이유가 장거리 수송을 위해서인가’라는 문제는 아직 단순하지 않다. 일반적인 배양 뉴런에서는 분열을 억제해 미토콘드리아를 길게 만들어도 축삭 유입이 크게 줄지 않았지만, 더 긴 축삭을 형성하도록 설계된 실험계에서는 작은 미토콘드리아가 더 풍부하고 분열/융합 단백질 비율도 달랐다. 즉 짧은 형태가 긴 축삭에서 특히 중요할 가능성은 있지만, 모든 조건에서 동일하게 적용되는 단순 법칙으로 보기에는 아직 이르다.
5. 품질관리는 어떻게 이루어지는가: 뉴런에서 mitophagy의 특별한 어려움
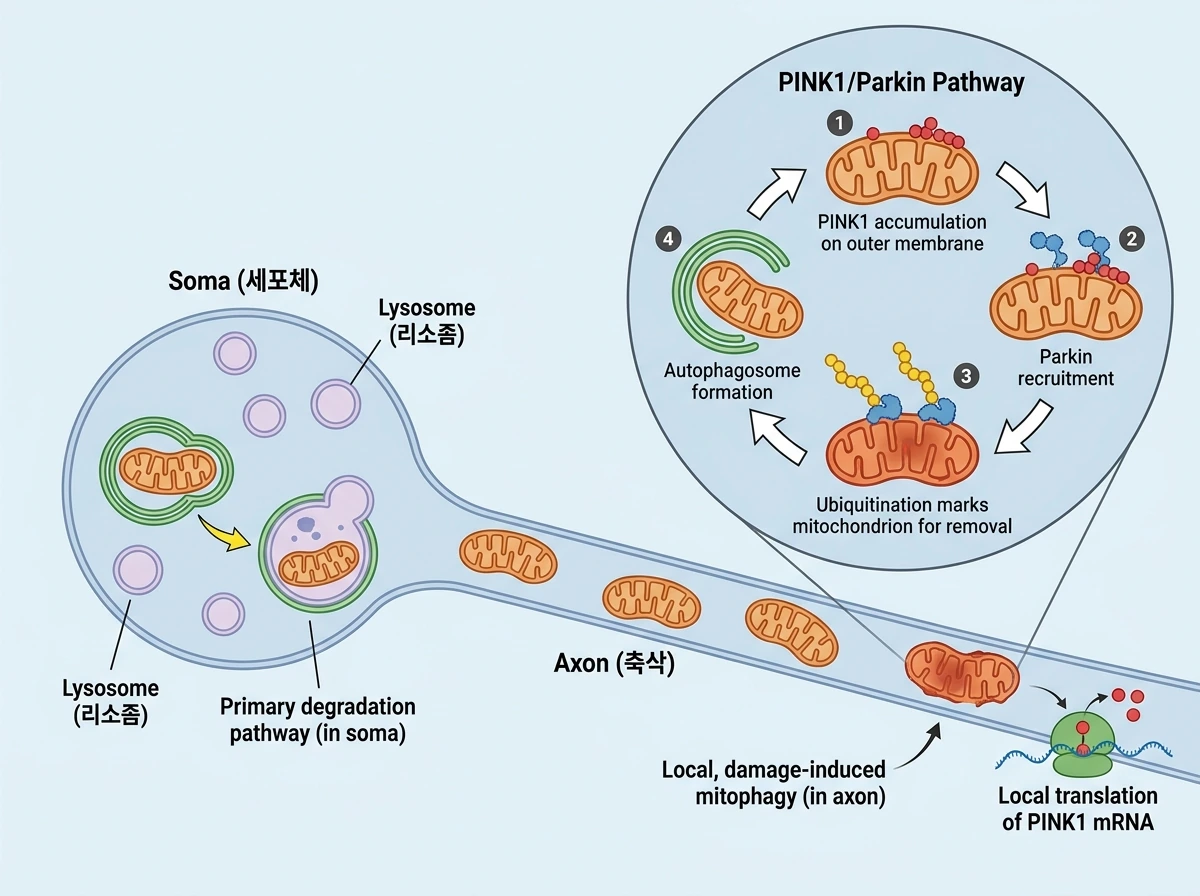
비분열성 세포인 뉴런에서는 미토콘드리아 수를 일정하게 유지하려면 생성과 제거의 균형이 중요하다. 이때 제거를 담당하는 대표 경로가 mitophagy이다. 그러나 리뷰에 따르면 뉴런의 basal mitophagy는 생각보다 낮다. 이는 손상된 미토콘드리아를 쉽게 제거하기보다, 충분히 심한 손상에만 반응하도록 임계값이 높게 설정되어 있기 때문일 수 있다. 저자들은 이것이 뉴런의 구조적 특성과도 맞물린다고 본다. 미토콘드리아 수송 속도는 빠르지 않고, 뇌 미토콘드리아 단백질의 반감기는 길기 때문에, 말단 부위의 미토콘드리아를 너무 쉽게 제거하면 오히려 현장 공급이 흔들릴 수 있다는 것이다. mitophagy가 어디서 일어나는가도 활발한 연구 주제다. 리소좀은 세포체와 수상돌기에 더 많으므로 기본적인 분해는 soma 쪽에서 활발할 가능성이 크다. 반면 축삭에서도 손상 유도형의 국소 mitophagy가 관찰되며, 축삭 말단이나 랑비에 결절 같은 부위에서도 관련 현상이 보고되었다. 따라서 현재의 그림은 ‘기본적인 분해는 soma에서 더 두드러지지만, 손상 상황에서는 축삭에서도 국소적으로 제거가 가능하다’는 다중 구획 모델에 가깝다. 가장 잘 알려진 분자 경로는 PINK1/Parkin 경로다. 건강한 미토콘드리아에서 빨리 분해되던 PINK1이 손상 시 바깥막에 축적되고, Parkin을 불러와 유비퀴틴화를 유도함으로써 자가포식 표지를 만든다. 이후 손상된 미토콘드리아 일부는 막 구조에 둘러싸여 autophagosome-lysosome 경로로 넘어간다. 또한 앞 절에서 본 분열과 품질관리는 서로 떨어진 현상이 아니라, 손상된 부분을 먼저 분리하고 그 일부를 선택적으로 제거하는 식으로 이어질 수 있다. 그런데 뉴런에서는 PINK1의 반감기가 짧아 먼 축삭 끝까지 단백질 자체를 운반하기 어렵다는 문제가 있다. 리뷰는 이 점을 뉴런 특이적 도전 과제로 강조하며, 실제로 포유류 뉴런에서 PINK1은 soma에서 운반될 뿐 아니라 축삭과 수상돌기에서 국소 번역된다고 설명한다. 이 덕분에 먼 부위에서도 mitophagy가 가능해진다는 것이다.
6. 미토콘드리아는 RNA 운반체이자 국소 번역의 거점이다
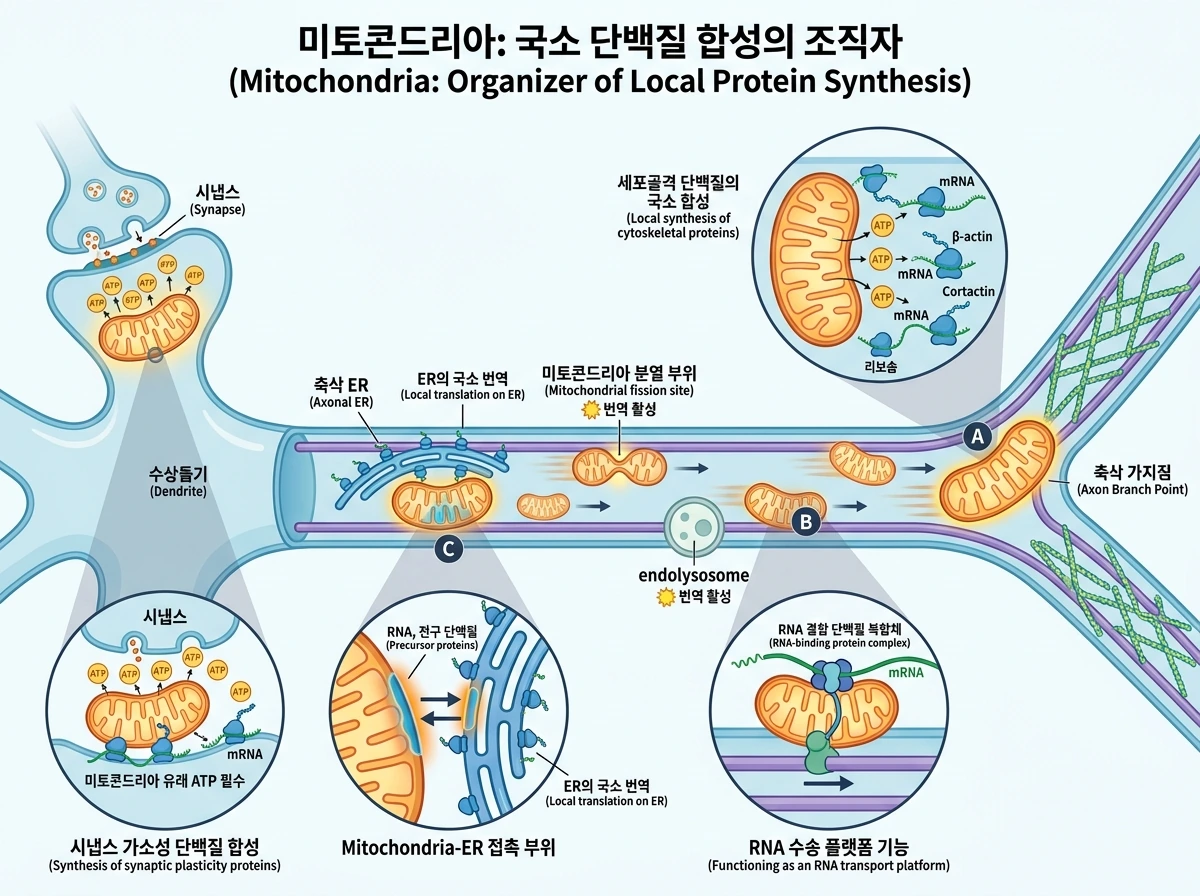
이 리뷰에서 특히 새롭게 강조되는 주제는 미토콘드리아가 국소 단백질 합성의 ‘발전소’일 뿐 아니라, 그 과정의 ‘조직자’일 수 있다는 점이다. 축삭에서는 번역이 미토콘드리아와 endolysosome의 접촉 부위, 미토콘드리아 분열 부위 주변, 그리고 축삭 가지 형성 부위에서 특히 활발하게 관찰된다. 이는 앞서 본 수송과 정박의 논리를 한 단계 확장한다. 즉 미토콘드리아가 어떤 구획에 도달해 멈춘다는 것은 단지 ATP를 소비 현장에 공급한다는 뜻이 아니라, 그 자리에서 새로운 단백질을 만드는 미세환경을 조성한다는 뜻이기도 하다. 리보솜은 endosome이나 ER와 연관되어 이동할 수도 있으며, 축삭 ER 역시 단순 구조물이 아니라 상당한 국소 번역 능력을 가진 것으로 제시된다. 이런 배경에서 미토콘드리아-ER 접촉 부위는 RNA와 전구 단백질의 이동 및 전달을 조율하는 자리로 해석된다. 일부 경우에는 RNA 결합 단백질 복합체와 mRNA가 미토콘드리아와 공간적으로 결합해 이동하는 모습도 제시되며, 이는 미토콘드리아가 번역에 필요한 분자를 태워 나르는 플랫폼처럼 기능할 수 있음을 시사한다. 중요한 것은 여기서 번역되는 단백질이 미토콘드리아 단백질만은 아니라는 점이다. 축삭 가지점에서는 베타-액틴과 cortactin 같은 세포골격 단백질의 국소 합성을 미토콘드리아가 뒷받침하고, 수상돌기에서는 시냅스 가소성에 필요한 단백질 합성에 미토콘드리아 유래 ATP가 필수적이다. 또한 앞 절의 PINK1 국소 번역은 이런 더 넓은 현상의 품질관리 버전으로 볼 수 있다. 즉 미토콘드리아는 단백질 합성이 일어날 ‘에너지적으로 유리한 장소’를 만들고, 경우에 따라 RNA 자체를 싣고 이동하는 플랫폼 역할도 수행한다.
7. OXPHOS 성분은 먼 축삭에서 어떻게 유지되는가: 합의와 가설
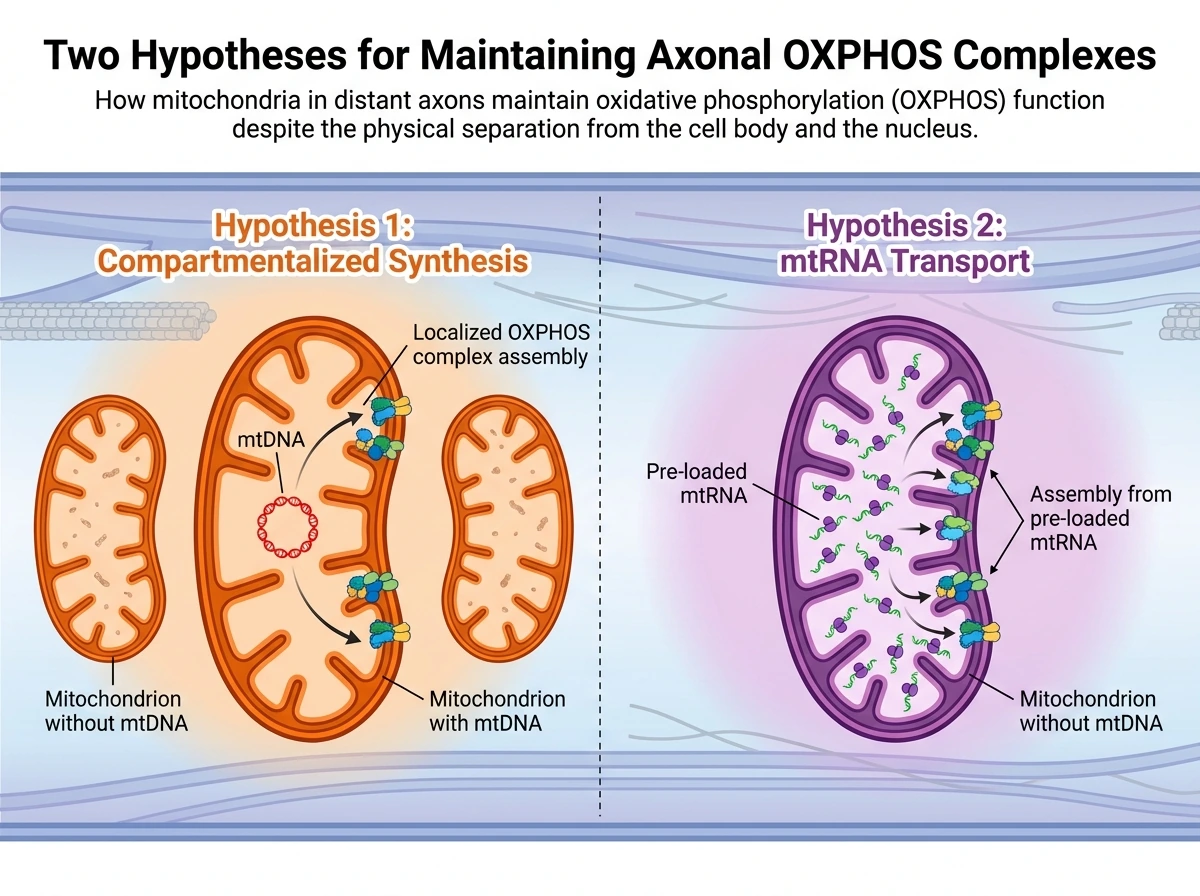
미토콘드리아와 국소 번역의 관계는 OXPHOS 복합체 생합성 문제에서 더욱 흥미로워진다. OXPHOS 복합체는 핵 DNA와 mtDNA가 각각 암호화한 단백질이 함께 조립되어야 완성된다. 리뷰는 시냅스 분획에서 국소적으로 합성된 OXPHOS 성분이 실제 복합체에 편입될 수 있고, 축삭 단백질 합성을 막으면 미토콘드리아 막전위가 떨어진다는 결과를 근거로, 먼 뉴런 구획에서도 OXPHOS 유지에 국소 번역이 필요하다고 본다. 이는 앞 절에서 본 ‘미토콘드리아-주변 번역 기계의 결합’이 단순한 보조 현상이 아니라, 에너지 생산 장치 자체의 유지 전략일 수 있음을 뜻한다. 또한 특정 번역 인자의 축삭 내 합성이 미토콘드리아 리보솜에 의한 mtDNA 유래 단백질 번역을 증가시킬 수 있다는 점도 소개된다. 그러나 여기서 아직 합의되지 않은 중요한 문제가 등장한다. 최근 연구에서는 일부 축삭 미토콘드리아가 mtDNA를 아예 갖지 않을 수 있다는 결과가 제시되었다. 만약 그렇다면 그 미토콘드리아는 현장에서 mtDNA 암호화 단백질을 새로 만들기 어렵다. 실제로 이런 미토콘드리아는 복합체 V를 역방향으로 작동시켜 ATP를 소모하면서도 막전위를 유지해 Ca2+ 유입 능력을 보존할 수 있다고 설명된다. 그렇다면 축삭에서 OXPHOS 복합체는 어떻게 유지될까? 리뷰는 이것을 아직 검증되지 않은 두 가지 가설로 제시한다. 하나는 mtDNA를 보유한 일부 미토콘드리아에 OXPHOS 생합성이 구획화되어 있을 가능성이다. 다른 하나는 축삭으로 이동하는 미토콘드리아 안에 mtDNA 대신 필요한 mtRNA가 충분히 실려 가는 시나리오다. 둘 다 가능성으로 남아 있으며, 저자들도 이 조율 메커니즘을 현재 활발한 연구 영역으로 규정한다. 이 대목은 이 리뷰가 확립된 지식과 가설적 모델을 분명히 구분하는 좋은 예다.
8. 뉴런 대사에서 미토콘드리아의 위치: ATP 생산만이 아니라 신경전달물질 대사의 허브
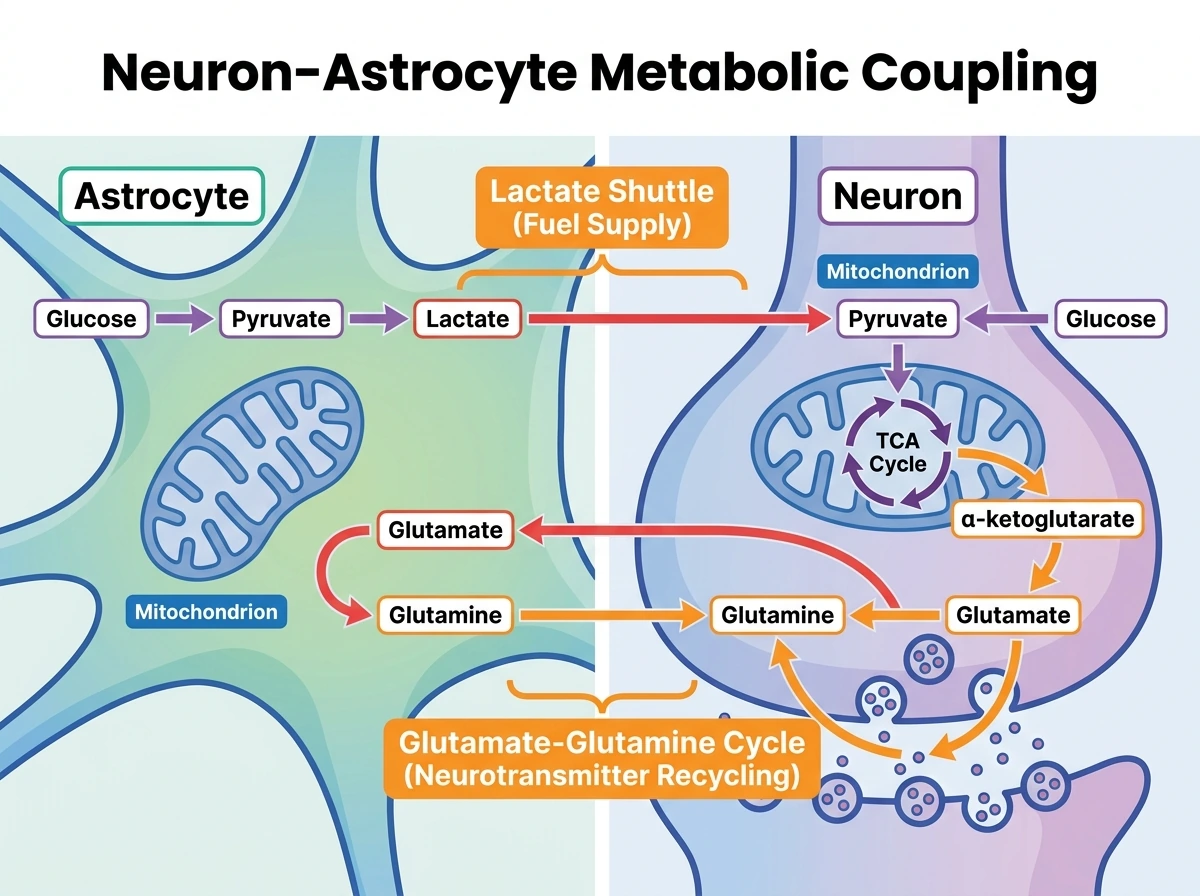
리뷰는 뉴런 미토콘드리아를 이해하려면 TCA 회로를 단순한 에너지 경로가 아니라 대사 허브로 봐야 한다고 강조한다. 앞선 절들이 주로 뉴런 내부의 공간 배치와 원거리 구획 유지에 초점을 두었다면, 여기서는 시야를 뉴런-아교세포 사이의 대사 교환으로 넓혀야 한다. 포도당에서 유래한 피루브산은 뉴런 자체의 해당과정을 통해 만들어져 미토콘드리아로 들어갈 수도 있고, astrocyte가 만들어 내보낸 lactate를 뉴런이 받아 다시 피루브산으로 바꾸어 사용할 수도 있다. 즉 탄소원은 세포질에서 바로 TCA 회로로 들어가는 것이 아니라, 바깥막과 안쪽막을 거쳐 미토콘드리아 기질로 유입된 뒤 산화대사에 편입된다. 이른바 lactate shuttle 모델은 뉴런과 astrocyte의 협력적 대사라는 관점을 제공한다. GABA와 glutamate 대사도 미토콘드리아와 깊게 연결된다. 뉴런 미토콘드리아의 TCA 중간체인 α-ketoglutarate는 glutamate와 연결되고, 이는 다시 억제성 뉴런에서 GABA 합성의 출발점이 된다. 시냅스 전달 후 astrocyte가 glutamate를 흡수해 glutamine으로 재생하고, 뉴런이 이를 다시 받아 glutamate/GABA 저장고를 보충하는 회로는 신경전달물질 순환이 곧 미토콘드리아 대사와 맞물려 있음을 보여 준다. 더 나아가 astrocyte 쪽 미토콘드리아도 glutamate와 GABA 처리에 직접 연결되어 있어, ‘뉴런은 에너지 생산만, astrocyte는 단순 회수만’이라는 식의 지나친 단순화는 피해야 한다. 현재의 더 적절한 정리는, 뉴런이 높은 전기생리학적 수요를 감당하기 위해 산화대사를 강하게 활용하는 한편, astrocyte와의 대사 교환이 신경전달물질 풀과 연료 공급을 함께 안정화한다는 것이다. 즉 미토콘드리아는 뉴런의 에너지 대사와 신경전달물질 순환, 그리고 세포 간 대사 분업이 만나는 접점이다.
9. 칼슘 처리: 뉴런 미토콘드리아가 특히 중요한 이유
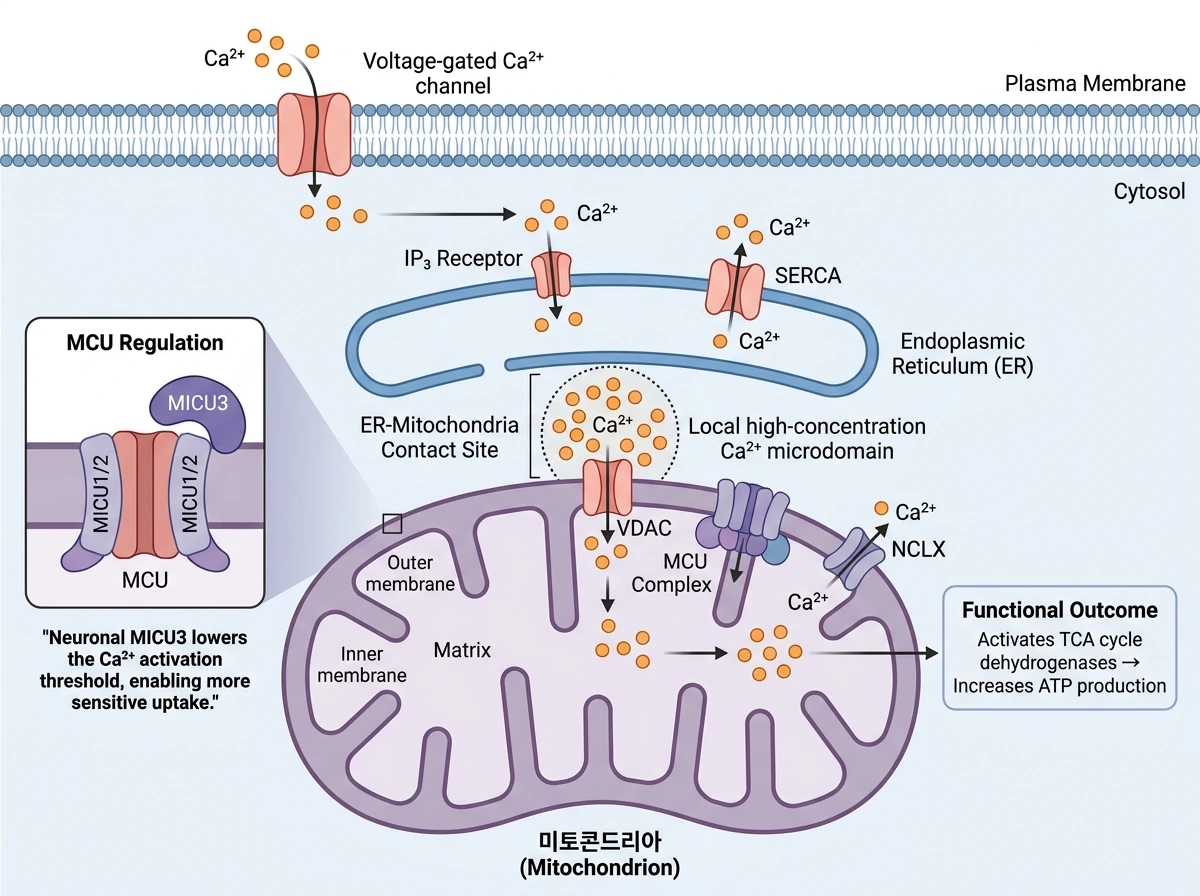
Ca2+ 조절은 이 리뷰의 또 다른 중심축이다. 신경세포는 활동할 때 세포질 Ca2+가 빠르게 오르내리므로, 이를 적절히 완충하면서 동시에 에너지 생산과 연결해야 한다. 미토콘드리아는 바로 이 지점에서 특별한 의미를 갖는다. Ca2+가 미토콘드리아 기질로 들어오면 TCA 회로의 주요 탈수소효소들이 활성화되어 OXPHOS와 ATP 생산이 증가한다. 즉 Ca2+ 유입 자체가 ‘지금 에너지가 더 필요하다’는 신호가 되는 셈이다. 이 과정은 세포질 전체의 평균 Ca2+가 아니라, 막 근처와 ER-미토콘드리아 접촉 부위에 형성되는 국소 고농도 미세영역에 크게 의존한다. 세포막의 전압의존성 Ca2+ 채널을 통해 들어온 Ca2+와, ER의 IP3 수용체나 ryanodine receptor를 통해 방출된 Ca2+가 이런 미세영역을 만들고, 미토콘드리아 바깥막의 VDAC와 안쪽막의 MCU가 이를 연속적으로 받아들인다. 반대로 SERCA는 Ca2+를 ER로 되돌려 보내므로, 뉴런의 Ca2+ 항상성은 세포막-ER-미토콘드리아가 연결된 회로로 보는 편이 이해하기 쉽다. 포유류에서 MICU1/2는 세포질 Ca2+가 충분히 높을 때 MCU를 열어 주는 조절자로 작동하고, 뉴런에 풍부한 MICU3는 그 작동 임계값을 더 낮춰 보다 민감한 Ca2+ 흡수를 가능하게 한다. 리뷰가 제시하는 비교에서는 MICU1/2 중심 조절보다 MICU3가 있을 때 더 낮은 Ca2+ 범위에서도 MCU 개방이 촉진될 수 있어, 뉴런 미토콘드리아가 일반 세포보다 활동 의존적 Ca2+ 신호를 더 예민하게 읽는다는 해석이 가능하다. 이는 뉴런 미토콘드리아가 일반 세포보다 빠르고 민감하게 활동 의존적 에너지 조절에 참여하도록 하는 특수화로 이해할 수 있다. 반대로 Ca2+ 방출은 주로 NCLX가 담당하고, Na+ 기울기가 무너지면 Ca2+/H+ antiporter가 대안이 될 수 있다. 병적 상황에서 MPTP가 오래 열리면 세포사멸로 이어질 수 있지만, 정상적인 Ca2+ 유출에 반드시 필요한 것은 아니라는 점도 구분해서 이해해야 한다. 최근에는 축삭과 수상돌기에서 MICU1/2와 NCLX 발현이 다를 수 있어 구획 특이적 Ca2+ 반응이 생긴다는 관찰도 소개되는데, 이는 뉴런 내부에서도 미토콘드리아 기능이 균일하지 않음을 시사한다.
10. 시냅스 기능과 가소성: ATP와 Ca2+가 만나는 자리
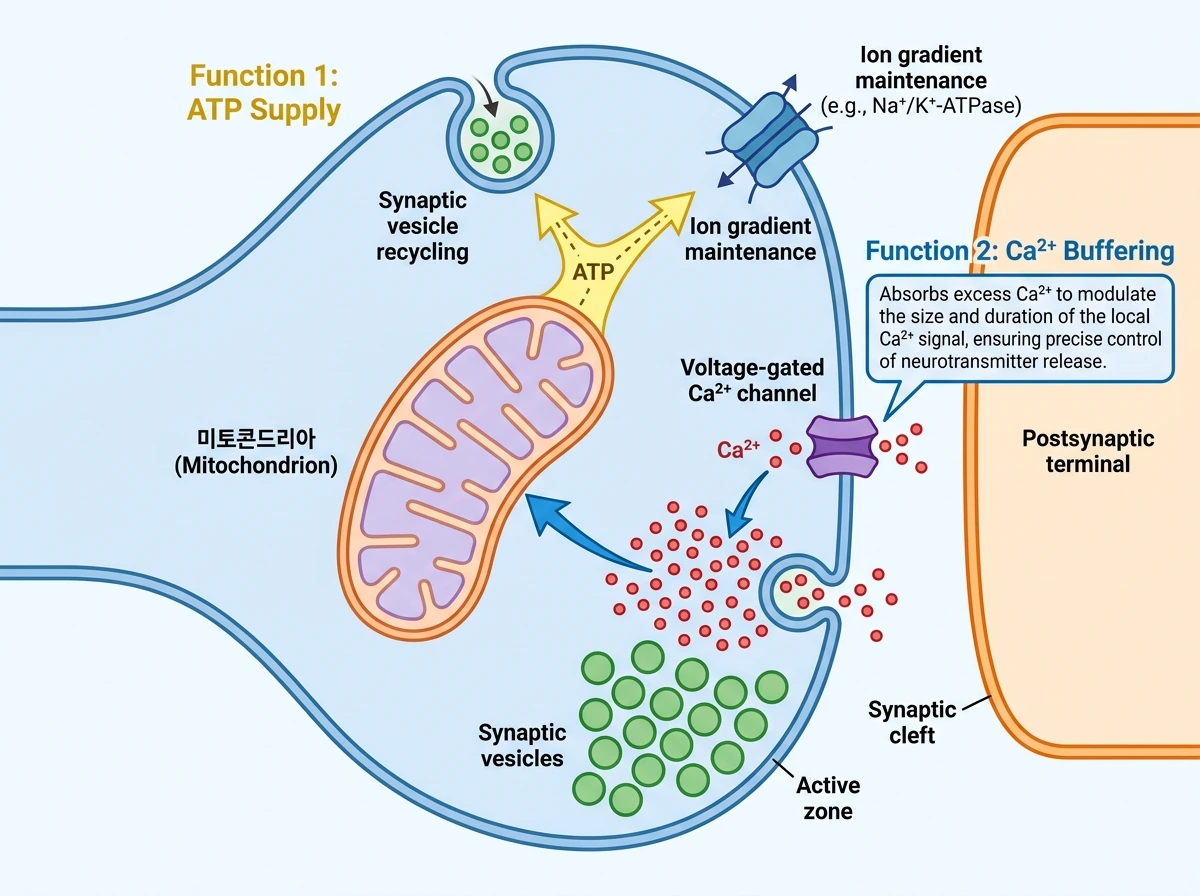
시냅스 수준에서 보면 미토콘드리아의 역할은 크게 두 갈래다. 첫째는 ATP 공급이다. 뉴런은 활동전위 후 막전위를 회복하는 Na+/K+-ATPase, 세포질 Ca2+를 제거하는 PMCA와 SERCA, 시냅스 소포 산성화를 위한 v-ATPase, 그리고 소포 재활용과 SNARE 분해·재조립 같은 과정에 막대한 ATP를 쓴다. ATP 공급이 흔들리면 이온기울기 복구가 느려지고, 신경전달이 약해지며, 흥분독성에 취약해진다. 도파민성 뉴런에서는 VMAT2가 도파민을 소포 안으로 격리해 세포질 산화를 막아 주는데, 이 역시 ATP 의존적인 소포 산성화 체계에 기대고 있어 미토콘드리아 기능과 연결된다. 둘째는 Ca2+ 완충이다. presynapse에서 미토콘드리아는 활동 시 막을 통해 들어온 Ca2+와 내부 저장고에서 나온 Ca2+의 일부를 흡수해, 활성 부위 주변 Ca2+ 미세영역의 크기와 지속시간을 바꾼다. 따라서 presynaptic mitochondrion은 단순히 ‘방출을 돕는 장치’라기보다, 방출의 시공간적 정밀도와 반복 자극 동안의 안정성을 조절하는 장치에 가깝다. postsynapse에서는 미토콘드리아 ATP와 Ca2+ 완충이 LTP와 LTD 같은 고전적 가소성 경로를 지지한다. 다만 여기서도 기전은 하나가 아니다. 미토콘드리아 분열은 적절한 LTP 유도에 필요하다고 제시되고, LTD 쪽은 cytochrome c 방출이나 caspase-3의 약한 활성화처럼 세포사멸 기계와 일부 겹치는 경로를 활용할 수 있다는 점이 논의된다. 즉 시냅스 가소성은 에너지 대사, Ca2+ 미세영역 조절, 형태 재편, 때로는 스트레스 신호 경로까지 겹쳐 나타나는 현상이며, 미토콘드리아는 그 교차점에 놓여 있다.
11. 질병과 연결되는 이유, 그리고 이 분야가 지금 중요한 이유
이 리뷰는 질병 자체를 깊게 다루지는 않지만, 왜 이 주제가 의학적으로 중요한지는 분명히 보여 준다. 미토콘드리아 기능 이상은 신경퇴행, 인지장애, 불안 관련 표현형 등과 연결되며, 특히 PINK1/Parkin 경로나 도파민 처리 과정과 같이 특정 취약성도 드러난다. 그러나 저자들이 강조하는 핵심은 ‘질병에서 망가진 미토콘드리아’를 보는 것만으로는 충분하지 않다는 점이다. 정상 뉴런에서 미토콘드리아가 어떻게 구획별로 다른 형태와 기능을 갖고, 어떻게 ATP 생산·Ca2+ 완충·산화환원 신호·국소 번역·품질관리를 조합하는지를 알아야, 질병에서 무엇이 실제로 무너졌는지도 정확히 이해할 수 있다. 특히 이 리뷰가 보여 주는 통합적 관점의 장점은, 수송 이상·대사 이상·칼슘 이상·자가포식 이상을 서로 다른 장으로 나누어 보는 대신 하나의 연결망으로 읽게 만든다는 데 있다. 또한 현재 기술적 한계도 분명하다. 살아 있는 뇌 안에서 작은 신경 구획별 미토콘드리아 활동을 충분한 시공간 해상도로 측정하기 어렵다. 그래서 고해상도 이미징, label-free 대사 측정, 대규모 omics, 그리고 이를 통합하는 기계학습 접근이 앞으로 분야를 크게 바꿀 것으로 전망된다. 즉 이 분야는 단순히 지식을 덧붙이는 단계가 아니라, 뉴런 생물학 전체를 다시 조직할 수 있는 전환점에 서 있다.
정리
이 리뷰가 제시하는 큰 메시지는 명확하다. 뉴런에서 미토콘드리아는 ATP 생산 소기관이라는 교과서적 정의를 넘어, 위치 선정, 이동과 정박, 분열과 융합, 품질관리, 국소 번역, Ca2+ 처리, 신경전달물질 대사, 그리고 뉴런-아교세포 대사 협력까지 통합하는 다기능 플랫폼이다. 이 기능들은 병렬적으로 나열되는 것이 아니라 서로 이어진다. 이동은 특정 구획에서의 ATP 공급과 Ca2+ 완충을 가능하게 하고, 그 위치 선택은 다시 국소 번역과 시냅스 안정성에 영향을 주며, 형태 변화는 수송 효율과 품질관리의 전제가 되고, 대사 상태와 칼슘 신호는 분열·정박·생존 결정에 되먹임을 준다. 현재 비교적 잘 확립된 내용은 미토콘드리아의 위치와 동역학이 뉴런 기능에 결정적이라는 점, ATP 생산과 Ca2+ 완충이 시냅스 전달과 가소성에 핵심이라는 점, 그리고 PINK1/Parkin 같은 품질관리 경로와 국소 번역이 먼 신경돌기에서 특히 중요하다는 점이다. 반면 아직 논쟁적이거나 미해결인 문제도 적지 않다. 왜 모든 presynapse가 미토콘드리아를 갖지 않는지, 구획별 형태 차이가 어떻게 만들어지는지, mtDNA가 없는 축삭 미토콘드리아가 어떻게 OXPHOS 기능을 유지하는지, 뉴런 종류마다 미토콘드리아 특수화가 얼마나 다른지가 대표적이다. 학부 수준에서 이 리뷰를 읽을 때 가장 중요한 포인트는 세부 단백질 이름을 외우는 것보다, ‘뉴런의 공간적 구조와 활동 패턴이 미토콘드리아의 기능적 특수화를 요구한다’는 큰 논리를 붙잡는 것이다. 그 틀을 이해하면, 미토콘드리아는 더 이상 배경 소기관이 아니라 신경세포 생리와 병리의 중심 요소로 보이기 시작한다.