
도입

발달 중인 뇌에서는 신경 회로가 올바르게 기능하기 위해 시냅스의 형성(formation)과 불필요한 시냅스의 제거(elimination/pruning)가 동시에 일어납니다. 중추신경계의 주요 교세포인 성상교세포(Astrocyte)는 시냅스 형성을 촉진하고, 미세아교세포(Microglia)는 시냅스를 포식하여 제거하는 역할을 담당합니다. 하지만 이 두 교세포가 어떻게 소통하여 시냅스 네트워크의 균형을 맞추는지는 오랫동안 풀리지 않은 의문이었습니다. 본 연구는 성상교세포가 분비하는 시냅스 형성 유도 단백질인 Hevin(SPARCL1)이 단백질 분해 효소에 의해 절단된 후, 미세아교세포의 TLR4 수용체에 결합하여 시냅스 제거를 촉진한다는 새로운 '교세포 간 신호전달(Glia-glia crosstalk)' 기전을 규명했습니다.
발달 과정에서의 시냅스 정제와 미세아교세포의 필수적 역할
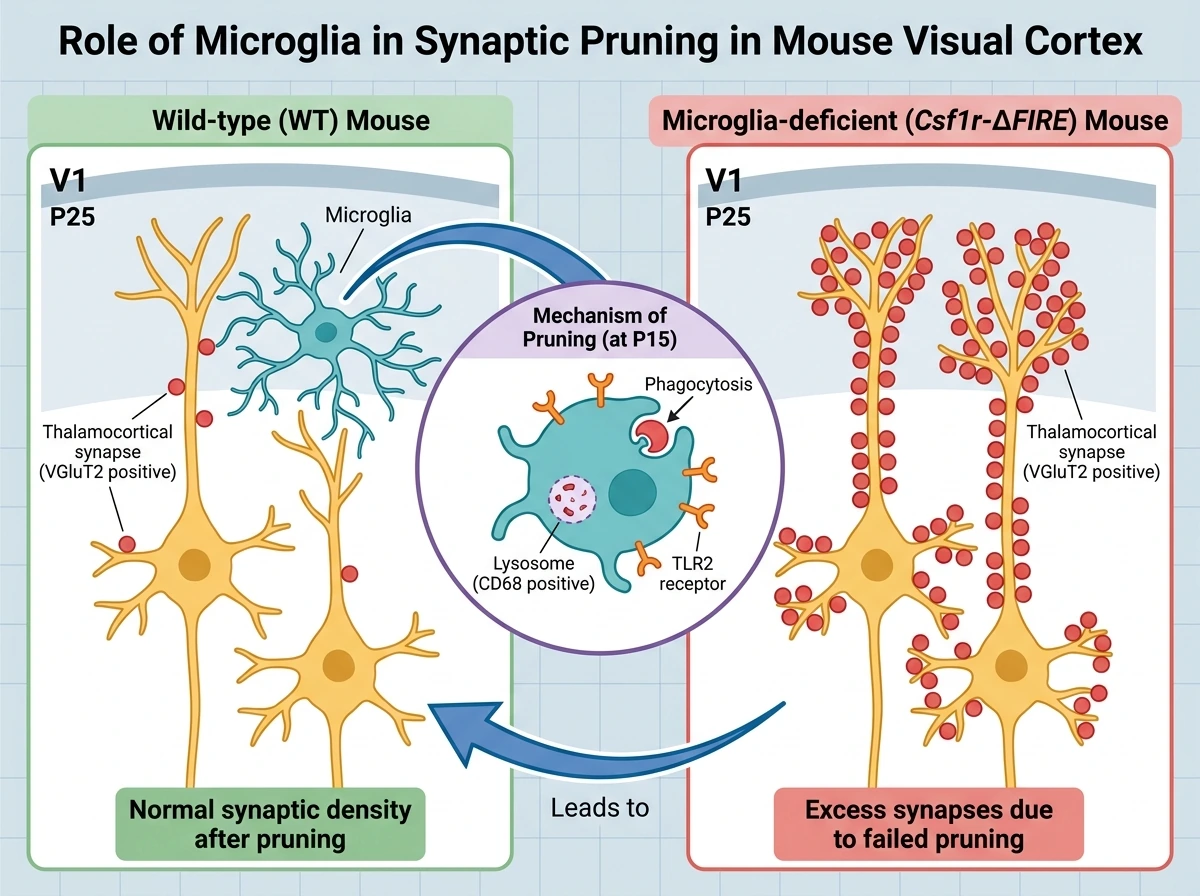
연구진은 생쥐의 일차시각피질(V1) 발달 과정을 관찰하여, 생후 10일(P10)에서 25일(P25) 사이에 피질 상층부(upper layers)를 중심으로 시상피질(thalamocortical) 시냅스의 밀도가 유의미하게 감소하는 시냅스 정제 현상을 확인했습니다. 이 시기, 특히 눈을 뜨는 시기 전후인 P15에 미세아교세포 내부에 리소좀 마커(CD68)가 증가하며 시냅스 전 말단(VGluT2)을 포식하는 현상이 활발해졌습니다. 흥미롭게도 발달 초기인 P10에는 미세아교세포에서 면역 수용체인 TLR2의 발현이 높게 나타났으나, 시냅스 정제가 진행되는 P15 이후에는 점차 감소하는 양상을 보였습니다. 미세아교세포가 시냅스 제거에 미치는 영향을 직접 확인하기 위해 미세아교세포가 결손된 생쥐 모델(Csf1r-ΔFIRE)을 분석한 결과, 야생형(WT)과 달리 P15와 P25 시기에 피질 상층부의 시냅스가 정상적으로 제거되지 않고 과도하게 남아있음을 발견했습니다. 이는 미세아교세포가 발달기 시각피질의 시냅스 정제에 필수적인 역할을 수행함을 명확히 보여줍니다.
Hevin의 절단과 미세아교세포로의 직접적인 신호 전달
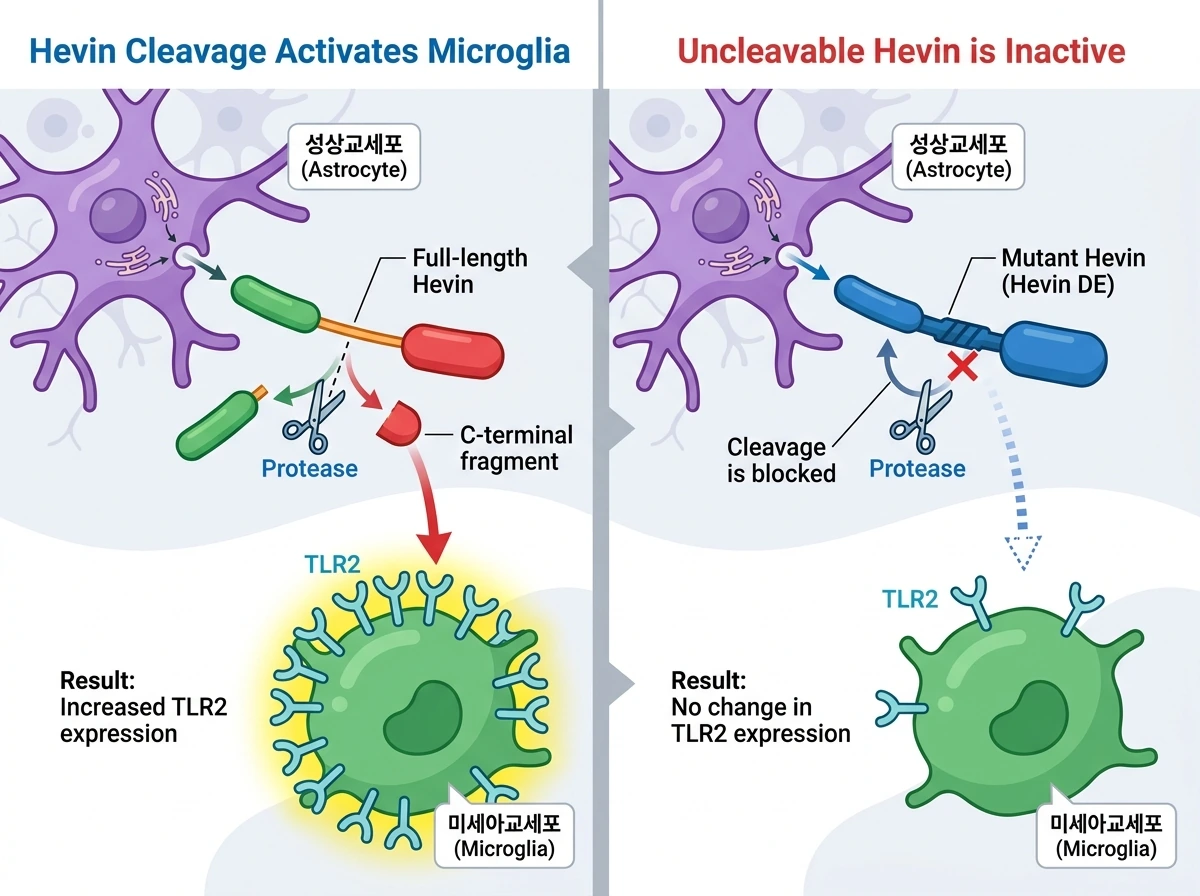
성상교세포가 분비하는 Hevin은 본래 시상피질 시냅스에 위치하여 시냅스 형성을 돕는 단백질입니다. 앞선 체외(in vitro) 실험에서 연구진은 분리된 미세아교세포에 Hevin을 처리했을 때, 염증을 유발하는 LPS와 달리 리소좀 및 세포골격 관련 유전자와 Tlr2 발현이 크게 증가함을 확인했습니다. 특히 단백질 분해 효소에 의해 절단된 'C-말단 조각(C-terminal fragment)'만이 이러한 반응을 유도했습니다. 이를 생체 내(in vivo) 환경에서 검증하기 위해, 바이러스 벡터(AAV)를 이용해 성상교세포에서 Hevin 또는 그 C-말단 조각을 과발현시킨 결과, 주변 미세아교세포에서 TLR2 발현이 뚜렷하게 증가했습니다. 반면, 효소에 의해 절단되지 않도록 유전적으로 조작한 돌연변이 Hevin(Hevin DE)을 발현시켰을 때는 대조군과 마찬가지로 TLR2 발현이 유도되지 않았습니다. 이는 Hevin이 미세아교세포에 신호를 전달하기 위해서는 단백질 분해 효소에 의한 절단 과정이 생체 내에서도 필수적임을 증명합니다.
Hevin이 유도하는 미세아교세포의 전사적 상태 변화와 포식 작용 촉진
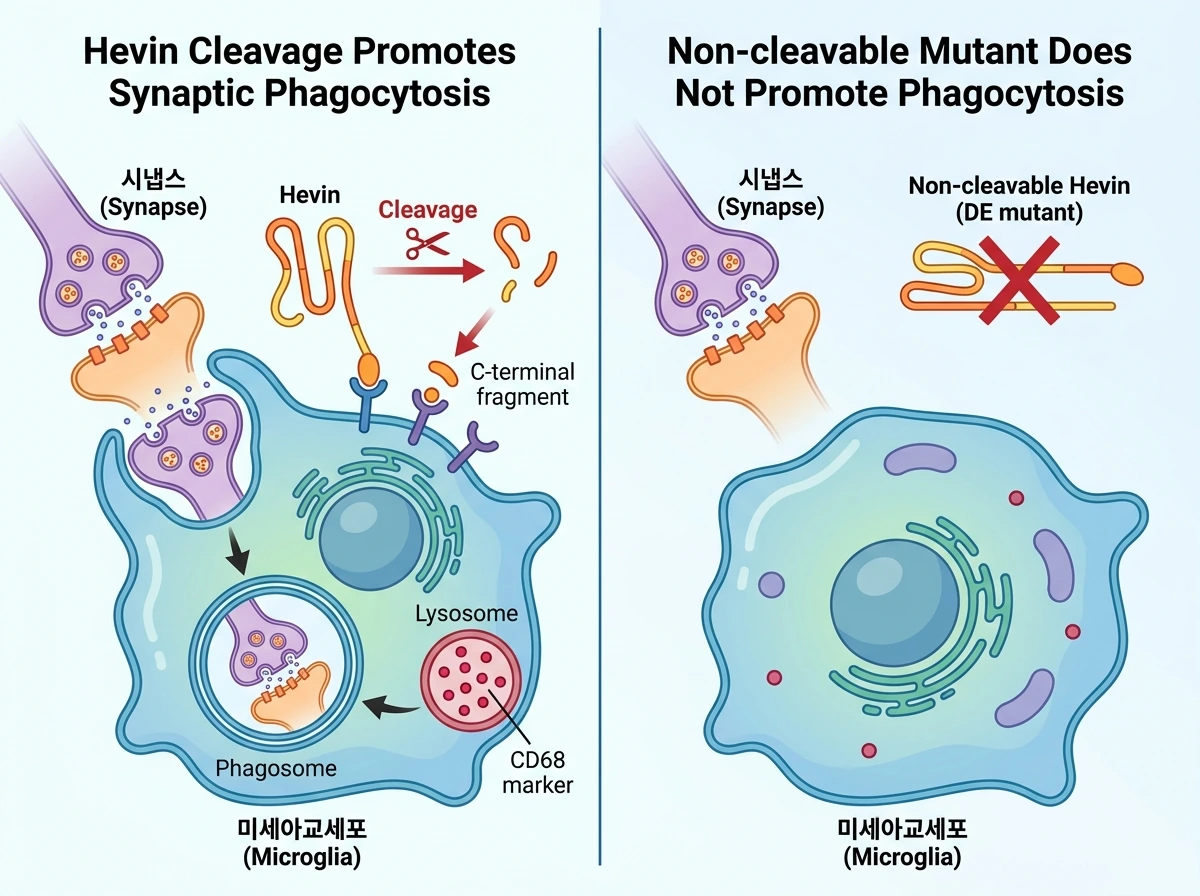
Hevin이 미세아교세포의 기능에 미치는 영향을 더 깊이 이해하기 위해 단일 세포 RNA 시퀀싱(scRNA-seq)을 수행한 결과, Hevin 과발현은 미세아교세포를 파고좀(Phagosome) 및 리소좀 관련 유전자(Apoe 등) 발현이 높은 특정 전사적 상태로 변화시켰습니다. 기존 데이터베이스와 비교 분석한 결과, 이 상태는 생후 4~5일경의 초기 발달 단계에서 관찰되는 포식성이 강한 미세아교세포(PAMs)나 백질 탈수초화 손상 시 나타나는 반응성 미세아교세포의 전사체 양상과 매우 유사했습니다. 실제로 조직 분석을 통해 확인해 보니, TLR2 발현이 높은 미세아교세포는 발현이 낮은 세포에 비해 리소좀 마커인 CD68을 유의하게 더 많이 함유하고 있었으며, 주변의 시상피질 시냅스를 리소좀 내부로 훨씬 더 활발하게 포식했습니다. 나아가, 온전한 Hevin이나 C-말단 조각을 과발현했을 때는 미세아교세포의 시냅스 포식이 크게 촉진되었지만, 절단되지 않는 돌연변이(Hevin DE)를 발현시켰을 때는 이러한 포식 작용 증가가 나타나지 않아 Hevin의 절단이 시냅스 제거 유도에 핵심적임을 다시 한번 확인했습니다.
TLR4 수용체를 매개로 한 시냅스 제거 메커니즘
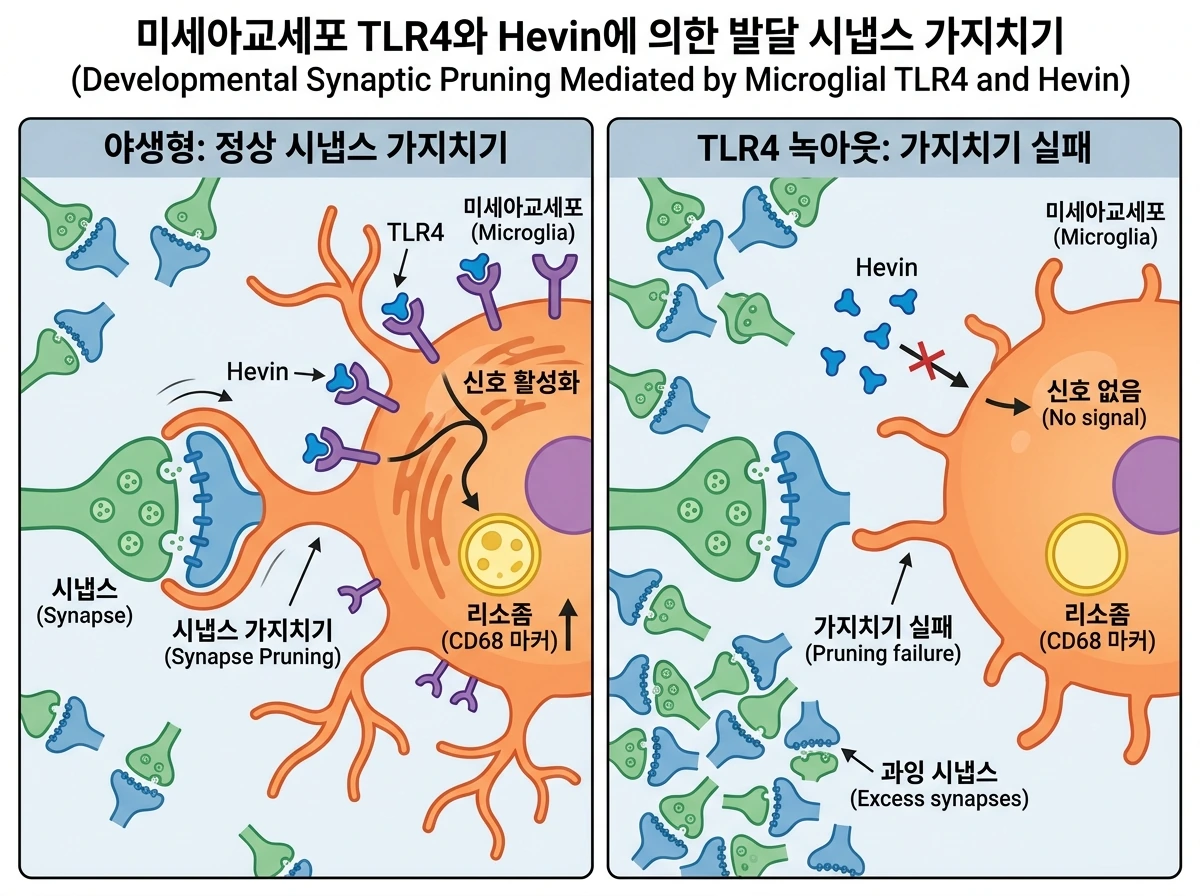
Hevin과 그 C-말단 조각이 미세아교세포에 신호를 전달하는 핵심 수용체는 선천성 면역 수용체로 잘 알려진 TLR4(Toll-like receptor 4)임이 확인되었습니다. 면역침강법(Co-IP) 분석 결과, Hevin은 미세아교세포 표면의 여러 수용체 중 TLR2나 MARCO가 아닌 TLR4와 특이적으로 결합했습니다. 또한 체외 실험에서 TLR 신호전달 억제제(OxPAPC)를 처리했을 때 Hevin에 의한 Tlr2 유전자 발현 증가가 완전히 차단되었습니다. 이를 생체 내에서 교차 검증하기 위해 TLR4가 결손된 생쥐(TLR4 KO)의 피질에 바이러스를 통해 Hevin을 과발현시킨 결과, 야생형 생쥐에서 관찰되었던 미세아교세포의 TLR2 발현 유도 및 리소좀 마커(CD68) 증가 현상이 전혀 나타나지 않았습니다. 나아가, TLR4 전체 결손 및 미세아교세포 특이적 조건부 결손 모델을 분석한 결과, 발달기 동안 제거되어야 할 시상피질 시냅스가 과도하게 남아있었고, 전기생리학적 기록(mEPSC)에서도 피질 뉴런의 흥분성 시냅스 활성이 비정상적으로 증가해 있음을 확인했습니다. 이는 미세아교세포의 TLR4 신호전달이 Hevin의 신호를 받아 발달기 시냅스 제거를 매개하는 데 필수적임을 의미합니다.
정리
본 연구는 성상교세포가 분비하는 Hevin이 온전한 상태에서는 시냅스를 안정화하지만, 효소에 의해 절단된 후에는 미세아교세포의 TLR4 수용체를 자극하여 시냅스 제거를 유도하는 '이중 역할'을 수행함을 규명했습니다. 이는 뇌 발달 과정에서 성상교세포와 미세아교세포가 어떻게 정교하게 소통하여 신경 회로의 균형을 맞추는지 보여주는 중요한 발견입니다. 나아가 이 교세포 간의 대화 메커니즘은 뇌 발달뿐만 아니라, 탈수초성 질환 등 신경 면역 질환에서 시냅스 및 세포 잔해 제거가 조절되는 과정을 이해하고 새로운 치료 표적을 발굴하는 데 중요한 기초 지식을 제공합니다.